09 July 2020: Lab/In Vitro Research
miR-512-3p Overcomes Resistance to Cisplatin in Retinoblastoma by Promoting Apoptosis Induced by Endoplasmic Reticulum Stress
Miao Kong1BCDEF, Yuanyuan Han2BCDEF, Yunhe Zhao3DF, Hong Zhang4AF*DOI: 10.12659/MSM.923817
Med Sci Monit 2020; 26:e923817






Abstract
BACKGROUND: Retinoblastoma (RB) seriously endangers the vision and even the life of patients. This study aimed to explore the endoplasmic reticulum stress (ERS) and drug resistance of RB and verify the effect of miR-512-3p as a regulator of XBP-1 shearing mechanism on ERS, proliferation, apoptosis, and autophagy levels of RB cells.
MATERIAL AND METHODS: Y79, weri-RB1, and HXO-RB44 cells were treated with cisplatin (DDP) gradient concentration to construct DDP-resistant cells. The drug inhibition rate and cell proliferation were assessed by CCK-8 assay. The cell transfection and cell apoptosis were detected by RT-qPCR analysis and TUNEL assay, respectively. The protein expression was detected by Western blot analysis. Dual-luciferase reporter assay confirmed the combination of miR-512-30p and XBP-1 transcript 1/2.
RESULTS: DDP inhibition rates for DDP-resistant RB cells were lower than that for RB cells. The XBP-1 expression was increased in DDP-resistant RB cells, and Y79 cells were chosen for the subsequent experiments. After transfection, miR-512-3p overexpression obviously inhibited the proliferation of DDP-resistant Y79 cells (Y79/DDP cells). miR-512-3p overexpression increased the DDP inhibition rate for Y79/DDP cells and apoptosis of Y79/DDP cells. miR-512-3p overexpression downregulated the expression of LC3 II/I in Y79/DDP cells. The effect of miR-512-3p inhibition on Y79/DDP cells was not as obvious as the effect of miR-512-3p overexpression on Y79/DDP cells. Furthermore, miR-512-3p was confirmed to be combined with XBP-1 transcript variant 1.
CONCLUSIONS: miR-512-3p improved the DDP resistance of RB cells by promoting ERS-induced apoptosis and inhibiting the proliferation and autophagy of RB cells.
Keywords: Endoplasmic reticulum stress, Retinoblastoma, Apoptosis Regulatory Proteins, Cisplatin, Retinal Neoplasms
Background
Retinoblastoma (RB) is the most common primary intraocular malignancy in children and is seen most frequently in infants. Approximately 90% of children with RB develop the disease before the age of 3 years. An investigation has found that the incidence of RB has been on the rise in recent years [1]. China has a high birth rate, with about 1100 new patients every year, of which 84% are advanced patients [2]. Although in recent years laser photocoagulation therapy, transpupillary thermotherapy, cryotherapy, selective arterial infusion chemotherapy, photodynamic therapy, and scleral applicator radiotherapy have been developed, the main treatment methods are still surgical removal of the eyeball and postoperative platinum-based chemotherapy [3,4]. The majority of RB children in remote parts of China almost all underwent enucleation or evisceration of the orbit once diagnosed; even so, the prognosis of RB was not improved [5]. According to statistics, the 5-year survival rate of RB patients with 1 eye was 50%, and that of binocular RB patients with only 1 eye removed was only 35% [6]. However, most of the children are not sensitive to chemotherapy, with poor prognosis and low long-term survival rate. Therefore, it is particularly important to find the target and mechanism of cisplatin resistance in RB.
More and more studies have found that endoplasmic reticulum stress (ERS), which activates the unfolded protein response (UPR), is involved in tumor development [7]. In tumor cells, ERS can improve homeostasis and make the surrounding environment suitable for tumor survival and growth [8]. Many negative factors such as hypoxia, nutrient deprivation, pH changes, or poor vascularization can hamper the growth of tumor cells, and thus activate the unfolded protein response (UPR) [9,10]. XBP-1 is an x-box binding protein. The XBP-1 gene is first transcribed into the unspliced XBP-1 (XBP-1u), and ERS can induce XBP-1u to be cut into spliced XBP-1 (XBP-1s) [11–13]. Under the condition of ERS, the mRNA of XBP-1u is cut into XBP-1s with high transcriptional activity, which promotes the UPR [14–17]. In a variety of tumors, XBP-1s expression is higher than in normal tissue [18–20]. Therefore, high expression of XBP-1s promotes the UPR, which is considered to be beneficial for cancer survival and development.
Studies have shown that ERS is related to the development of RB [16-18]. Multiple molecular biological information databases reveal that miR-512-3p has a potential binding site (inhibiting XBP-1u) that regulates its mRNA splicing. miR-512-3p was also investigated in prostate cancer [21], non-small cell lung cancer [22], and hepatocellular carcinoma [23], and miR-512-3p overexpression suppressed the cell proliferation and tumor invasion ability in these cancers. The role of miR-512-3p in retinoblastoma remains to be studied.
In this study, we explored the ERS and drug resistance of RB and assessed the effect of miR-512-3p as a regulator of XBP-1 shearing mechanism in ERS, as well as proliferation, apoptosis, and autophagy levels of RB cells.
Material and Methods
CONSTRUCTION OF CISPLATIN (DDP)-RESISTANT CELLS:
Y79 cells were obtained from American Type Culture Collection (Rockville, MD, USA). weri-RB1 and HXO-RB44 cells were brought from Shanghai Zishi Biotechnology Co. These cells were cultured in RPMI-1640 medium with a final concentration of 20% fetal bovine serum (FBS) at 37°C with an atmosphere of 95% O2 and 5% CO2. Cells were seeded in the 96-well plate with approximately 8000 cells in each well, and were cultured for 2 h. Cells were divided into a blank group, a control group, and a treatment group. Cells in the treatment group were treated with 10 μL DDP at concentrations of 0.1, 1, 5, 10, 25, 50, and 100 μg/mL for 72 h. Blank groups were not added with DDP and cells and control groups were only added with cells without DDP.
CCK-8 ASSAY:
Cells in logarithmic growth stage were counted and inoculated in the 96-well plate for 24 h. After 24 h, 10 μL CCK8 solution was added into each well for another 4-h incubation of cells with oscillation. The absorbance (OD) value was measured at the wavelength of 490 nm by enzyme-linked immunoassay, and the OD value was zeroed in blank groups. Inhibition rate=[1–(ODtreatment–ODblank)/(ODcontrol–ODblank)]×100%.
WESTERN BLOT ANALYSIS:
RIPA buffer lysed the cells to obtain the total protein and a BCA kit was used to quantify the total protein concentration in each group of cells. We separated 30 μg proteins with SDS-PAGE and transferred them to PVDF membranes with semi-dry transfer method. We sealed the PVDF membrane with 50 g/L skim milk powder for 2 h at room temperature. The PVDF membrane was incubated with primary antibody at 4°C overnight, and with secondary antibody at 37°C for 1 h. GAPDH was used as an internal control for the detection of XBP-1 (ab37152, Abcam), LC3I/II (ab62721, Abcam), GRP78 (ab21685, Abcam), ATF4 (ab184909, Abcam), ATF6 (ab203119, Abcam), IRE1 (ab37073, Abcam), CHOP (#2895, Cell Signaling Technology), XBP-1s (#27901, Cell Signaling Technology), XBP-1u (25997-1-AP, ProteinTech), Casp-4 (ab25898, Abcam), casp-3 (ab13847, Abcam), c-casp-3 (ab2302, Abcam), Bax (ab32503, Abcam), Bcl-2 (ab32124, Abcam), Beclin-1 (ab207612, Abcam) and Atg5 (#2630, Cell Signaling Technology). Then, ImageJ software was used for grayscale scanning and quantification.
CELL TRANSFECTION:
Cells in the logarithmic growth phase were taken and transferred to serum-free 1640 medium 2 h before transfection. Cells were divided into a control group, a mimic NC (miR1N0000001-1-5, Guangzhou Ruibo Biotechnology Co.) group, a miR-512-3p mimic (miR10002823-1-5, Guangzhou Ruibo Biotechnology Co.) group, an inhibitor NC (miR2N0000001-1-5, Guangzhou Ruibo Biotechnology Co.) group, and an miR-512-3p inhibitor (miR20002823-1-5, Guangzhou Ruibo Biotechnology Co.) group. After transfection, the cells were cultured at 37°C in incubators containing 5% CO2. After 4–6 h, the mixture was sucked out and replaced with normal medium for further cell culture for 24 h at 37°C with 5% CO2. RT-qPCR analysis was used to assess the transfection status of cells in each group. The procedures were conducted in strict accordance with the instructions of the kit.
RT-QPCR ANALYSIS:
After cell transfection, RNA was extracted with TRIzol reagent (No. 12183555, Thermo Fisher Scientific, Inc.). We prepared 10 μL total RNA taken for the reaction system for reverse transcription using a PrimeScript Reverse Transcription Reagent kit (No. RR037A, Takara). Next, the reaction system was prepared again, and the well-uploaded 8 tubulars were placed on the RT-qPCR instrument for reaction performed using a TaqMan Universal PCR Master Mix kit (No. 4364338, Thermo Fisher Scientific, Inc.) at 95°C for 3 min of pre-degeneration and 95°C for 12 s, and 62°C for 40 s for 40 cycles. The primer sequences for qPCR were:
TUNEL ASSAY:
The experiment was conducted using a TUNEL detection kit (Beijing ZhongShan Biotechnology Company). The main steps were: the sample slide was digested by protease K and then treated with TdT and Biotin-dUTP. After the sample slide was sealed by the sealing liquid, streptavidin-HRP working liquid and DAB color reagent were added. The color was observed and cells were counted under a light microscope.
DUAL-LUCIFERASE REPORTER ASSAY:
Analysis using ENCORI software predicted that miR-512-3p can combine with XBP-1, which was confirmed by dual-luciferase reporter assay. Cells were co-transfected with XBP-1 transcript variant 1 (u) and miR-512-3p mimic or mimic control (NC) vector, and cells were co-transfected with XBP-1 transcript variant 1 (s) and miR-512-3p mimic or mimic control (NC) vector. Lipofectamine® 2000 reagent was the transfection reagent. Luciferase activity was detected by a dual-luciferase reporter assay system with the Dual-Luciferase Assay Kit (Promega).
STATISTICAL ANALYSIS:
GraphPad Prism 6 Software was used to analyze data. Data are expressed as mean ± standard deviation. The Bonferroni
Results
DDP-RESISTANT CELLS WERE CONSTRUCTED BY GRADIENT CONCENTRATION OF DDP:
Y79, weri-RB1, and HXO-RB44 cells were treated with gradient concentration of DDP for 72 h. The DDP inhibition rates for DDP-resistant RB cells were decreased, especially for Y79/DDP cells (Figure 1A). The expression of XBP-1 in DDP-resistant RB cells was higher than that in RB cells (Figure 1B). Y79 cells were selected for subsequent experiments considering the induction effect of drug resistance and the expression of XBP-1.
DDP-RESISTANT Y79 CELLS (Y79/DDP CELLS) WERE TRANSFECTED:
Y79/DDP cells were transfected with mimic NC, miR-512-3p mimic, inhibitor NC, and miR-512-3p inhibitor. As shown in Figure 2, miR-512-3p expression was upregulated in Y79/DDP cells transfected with miR-512-3p mimic and was downregulated in Y79/DDP cells transfected with miR-512-3p inhibitor compared with the control group, mimic NC group, and inhibitor NC group.
PROLIFERATION OF Y79/DDP CELLS AND DDP INHIBITION RATE FOR Y79/DDP CELLS WERE CHANGED AFTER TRANSFECTION:
After transfection, miR-512-3p overexpression or inhibition all decreased the proliferation of Y79/DDP cells (Figure 3A). As shown in Figure 3B, miR-512-3p overexpression or inhibition increased the DDP inhibition rate of Y79/DDP cells. However, the effect of miR-512-3p inhibition on Y79/DDP cells was not as obvious as the effect of miR-512-3p overexpression on Y79/DDP cells.
MIR-512-3P AFFECTS THE APOPTOSIS AND AUTOPHAGY OF Y79/DDP CELLS:
As shown in Figure 4A, miR-512-3p overexpression or inhibition promoted the apoptosis of Y79/DDP cells, and the promotion effect of miR-512-3p overexpression on cell apoptosis was far stronger than that of miR-512-3p inhibition. The expression of LC3 II/I in Y79/DDP cells transfected with miR-512-3p mimic or inhibitor was decreased, and the decreased expression of LC3 II/I in the former cells was more obvious (Figure 4B).
MIR-512-3P AFFECTS THE RELATED PROTEINS EXPRESSION IN ERS:
The related proteins expression in ERS was determined by Western blot analysis. As shown in Figure 5A, miR-512-3p overexpression enhanced the expression of GRP78, ATF4, ATF6, and IRE1, while miR-512-3p inhibition suppressed the expression of GRP78, ATF4, ATF6, and IRE1 compared with that in Y79/DDP cells. As shown in Figure 5B, miR-512-3p overexpression increased the expression of CHOP and XBP-1s but was restrained by miR-512-3p inhibition. miR-512-3p overexpression also increased the Casp-4 expression, but was not obviously affected by miR-512-3p inhibition. However, miR-512-3p overexpression suppressed the XBP-1u expression, and miR-512-3p inhibition enhanced the XBP-1u expression.
MIR-512-3P AFFECTS THE RELATED PROTEINS EXPRESSION IN APOPTOSIS AND AUTOPHAGY:
The related proteins expression in apoptosis and autophagy was also detected by Western blot analysis. The result of Figure 6 indicated that miR-512-3p overexpression or inhibition upregulated the expression of c-casp-3 and Bax, but downregulated the Bcl-2 expression, and the role of miR-512-3p overexpression in apoptosis and autophagy was more obvious.
MIR-512-3P IS COMBINED WITH XBP-1 TRANSCRIPT VARIANT 1:
Bioinformatics analysis was performed, and the results of ENCORI predicted that miR-512-3p combined with XBP-1 (Figure 7A). The dual-luciferase reporter assay indicated that the relative luciferase activity was significantly decreased when cells were co-transfected with miR-512-3p mimic and XBP-1 transcript variant 1, but were not obviously changed when cells were co-transfected with miR-512-3p mimic and XBP-1 transcript variant 2 (Figure 7B). Importantly, miR-512-3p could combine with XBP-1 transcript variant 1.
Discussion
We explored the effect of miR-512-3p as a regulator of XBP-1 shearing mechanism on ERS, proliferation, apoptosis and autophagy levels of RB cells. We found that miR-512-3p could overcome the DDP resistance in RB cells by promoting the ERS to induce apoptosis and inhibit proliferation and autophagy of RB cells.
The endoplasmic reticulum (ER) is involved in the luminal chaperones for folding and packing of secretory proteins, intracellular trafficking, mitochondrial biogenesis, autophagy, apoptosis, and inflammation [24–26]. UPR is an evolutionally conserved cellular-protective response, which is the response of cells to ERS [27]. In cancer cells, stress conditions causing UPR are ubiquitous, such as the lack of oxygen and nutrients in the microenvironment of the tumor, which lead to increased dependence of cancer cells on UPR [28]. Studies have shown that UPR can induce drug resistance in tumor cells and reduce the efficacy of anti-tumor drugs [29–31]. High expression of XBP-1s can promote the development of cancer by promoting the UPR [14–17]. miR-512-3p can regulate the expression of XBP-1s and XBP-1u. In this study, we found that miR-512-3p inhibited proliferation and autophagy, promoted apoptosis, and overcame DDP resistance of RB cells by regulating the UPR affected by the expression of XBP-1s and XBP-1u. miR-512-3p has been demonstrated to a tumor-suppressive miRNA in some cancers [21–23]. The suppressive role of miR-512-3p in tumors was also demonstrated in this study.
UPR consists of 3 parallel signaling pathways. The main regulators of these pathways are 3 ER transmembrane proteins: PERK, IRE1, and ATF6. When ERS occurs in cells, it not only induces CHOP expression through ATF4, but also further activates the signaling pathways of 2 other ER transmembrane proteins – IRE1 and ATF6 – and subsequently activates PERK and eIF2. GRP78 can regulate the expression of PERK, IRE1, and ATF6 to participate in the transfer of unfolded proteins. Therefore, the induction of GRP78 is a recognized marker of ERS and UPR activation.
Studies have shown that inhibiting the ATF4 pathway in ERs can effectively reduce the proliferation and autophagy of tumors and improve the level of apoptosis [32–34]. CHOP is a direct target of ATF4, representing the pro-apoptotic component of UPR. After injection of tunicamycin, wild-type mice showed higher levels of renal epithelial cell apoptosis than CHOP knockout mice [35]. This study indicated that miR-512-3p overexpression promoted the expression of GRP78, ATF4, ATF6, IRE1, and CHOP, which was reversed by miR-512-3p inhibition. However, miR-512-3p overexpression or inhibition were not conducive to the health of RB cells, and the negative effects of miR-512-3p overexpression on RB cells were more significant.
Autophagy is considered to be an important mechanism to deal with ERS. Autophagy is a way to reduce ERS and allow cells to survive, which was found only in cancer cells, not in non-transforming cells [36–38]. In tumors, ERS-induced autophagy can also lead to tumor resistance to anti-tumor drugs. ERS can induce autophagy activation and make cells resistant to cisplatin-induced apoptosis [39]. In this study, enhanced or alleviated autophagy decreased the DDP inhibition rates of DDP/Y79 cells, and the effect of enhanced autophagy on DDP resistance was more obvious.
Conclusions
Inhibition rates in DDP-resistant RB cells were decreased and XBP-1 expression in DDP-resistant RB cells was increased. miR-512-3p increased the DDP inhibition rates for DDP/Y79 cells. miR-512-3p suppressed the proliferation and autophagy and promoted the apoptosis of DDP/Y79 cells by regulating the expression of XBP-1s and XBP-1u. The effects of miR-512-3p overexpression on DDP/Y79 cells were more obvious than that of miR-512-3p inhibition. Therefore, miR-512-3p overcame the DDP resistance of RB cells targeting XBP-1u by promoting ERS-induced apoptosis and inhibiting the proliferation and autophagy of RB cells. A limitation of our study is that it was a primary study and only 1 type of cell was studied throughout the whole experiment.
Figures
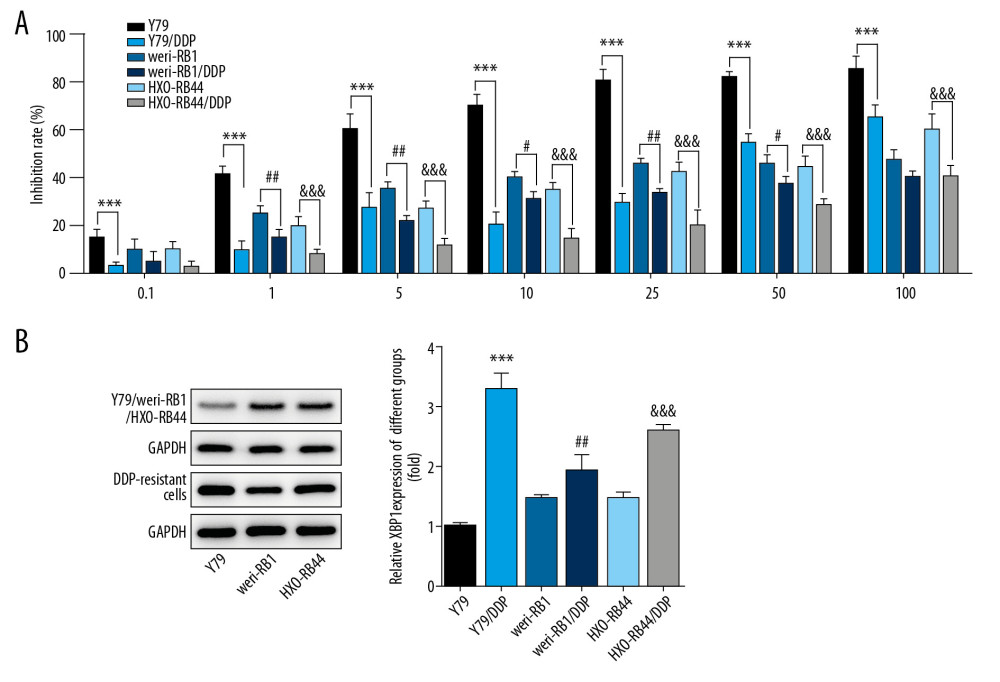 Figure 1. DDP-resistant cells were constructed by gradient concentration of DDP. (A) The DDP inhibition rates for DDP-resistant RB cells were reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group. (B) The expression of XBP-1 in DDP-resistant RB cells was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group.
Figure 1. DDP-resistant cells were constructed by gradient concentration of DDP. (A) The DDP inhibition rates for DDP-resistant RB cells were reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group. (B) The expression of XBP-1 in DDP-resistant RB cells was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group. 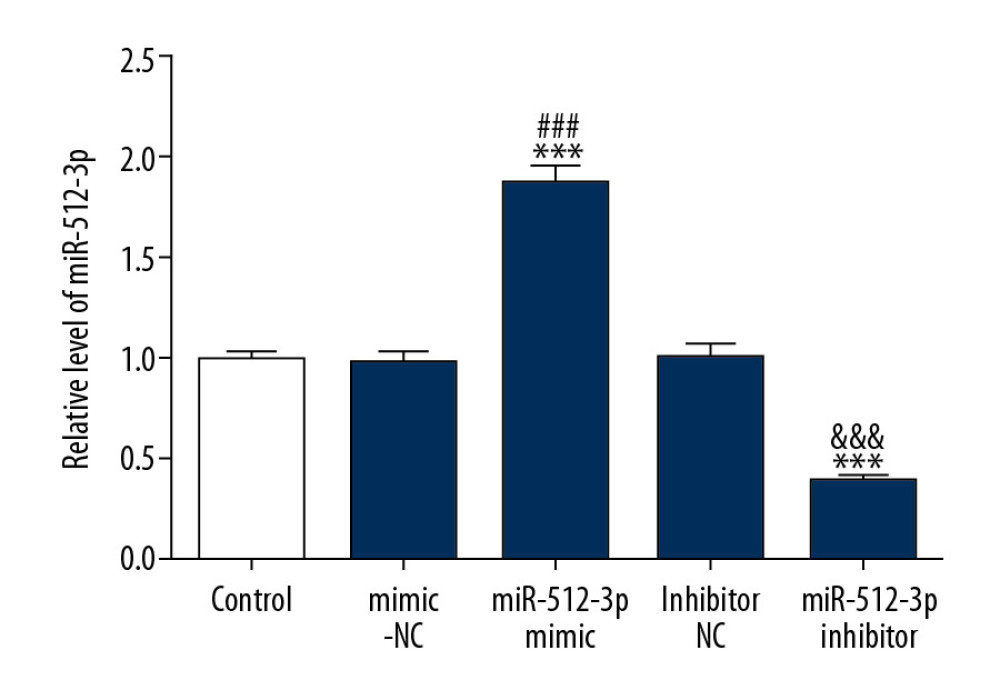 Figure 2. DDP-resistant Y79 cells (Y79/DDP cells) were transfected. RT-PCR analysis confirmed the transfection effects. *** P<0.001 vs. control group. ### P<0.001 vs. mimic NC group. &&& P<0.01 vs. inhibitor NC group.
Figure 2. DDP-resistant Y79 cells (Y79/DDP cells) were transfected. RT-PCR analysis confirmed the transfection effects. *** P<0.001 vs. control group. ### P<0.001 vs. mimic NC group. &&& P<0.01 vs. inhibitor NC group. 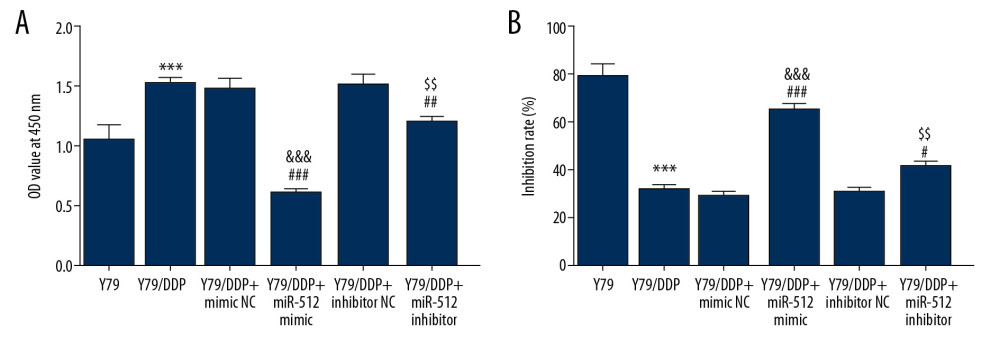 Figure 3. Proliferation of Y79/DDP cells and DDP inhibition rate for Y79/DDP cells were changed after transfection. (A) The proliferation of Y79/DDP cells after transfection was detected by CCK-8 assay. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group. (B) The DDP inhibition rates for Y79/DDP cells after transfection were also reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group.
Figure 3. Proliferation of Y79/DDP cells and DDP inhibition rate for Y79/DDP cells were changed after transfection. (A) The proliferation of Y79/DDP cells after transfection was detected by CCK-8 assay. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group. (B) The DDP inhibition rates for Y79/DDP cells after transfection were also reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group. 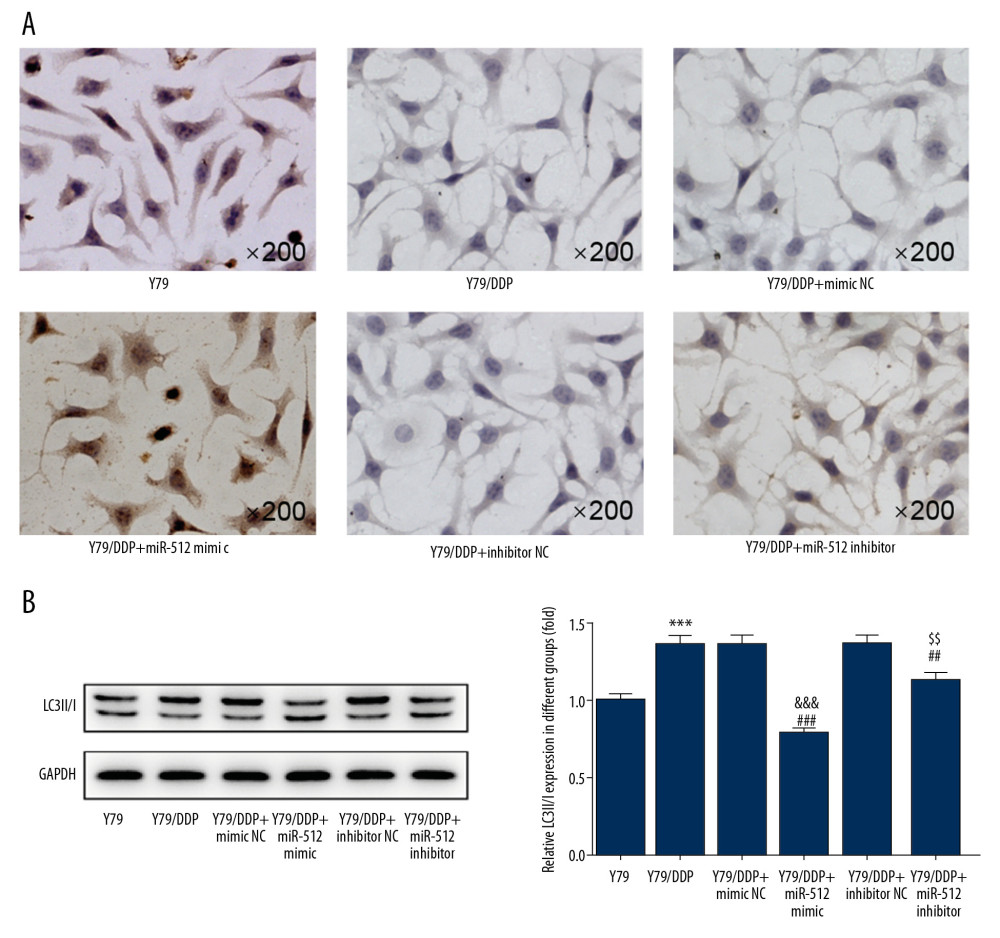 Figure 4. miR-512-3p affects the apoptosis and autophagy of Y79/DDP cells. (A) The apoptosis of Y79/DDP cells after transfection was determined by TUNEL assay. (B) The proteins expression of autophagy in Y79/DDP cells after transfection was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. ΔΔΔ P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group.
Figure 4. miR-512-3p affects the apoptosis and autophagy of Y79/DDP cells. (A) The apoptosis of Y79/DDP cells after transfection was determined by TUNEL assay. (B) The proteins expression of autophagy in Y79/DDP cells after transfection was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. ΔΔΔ P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group. 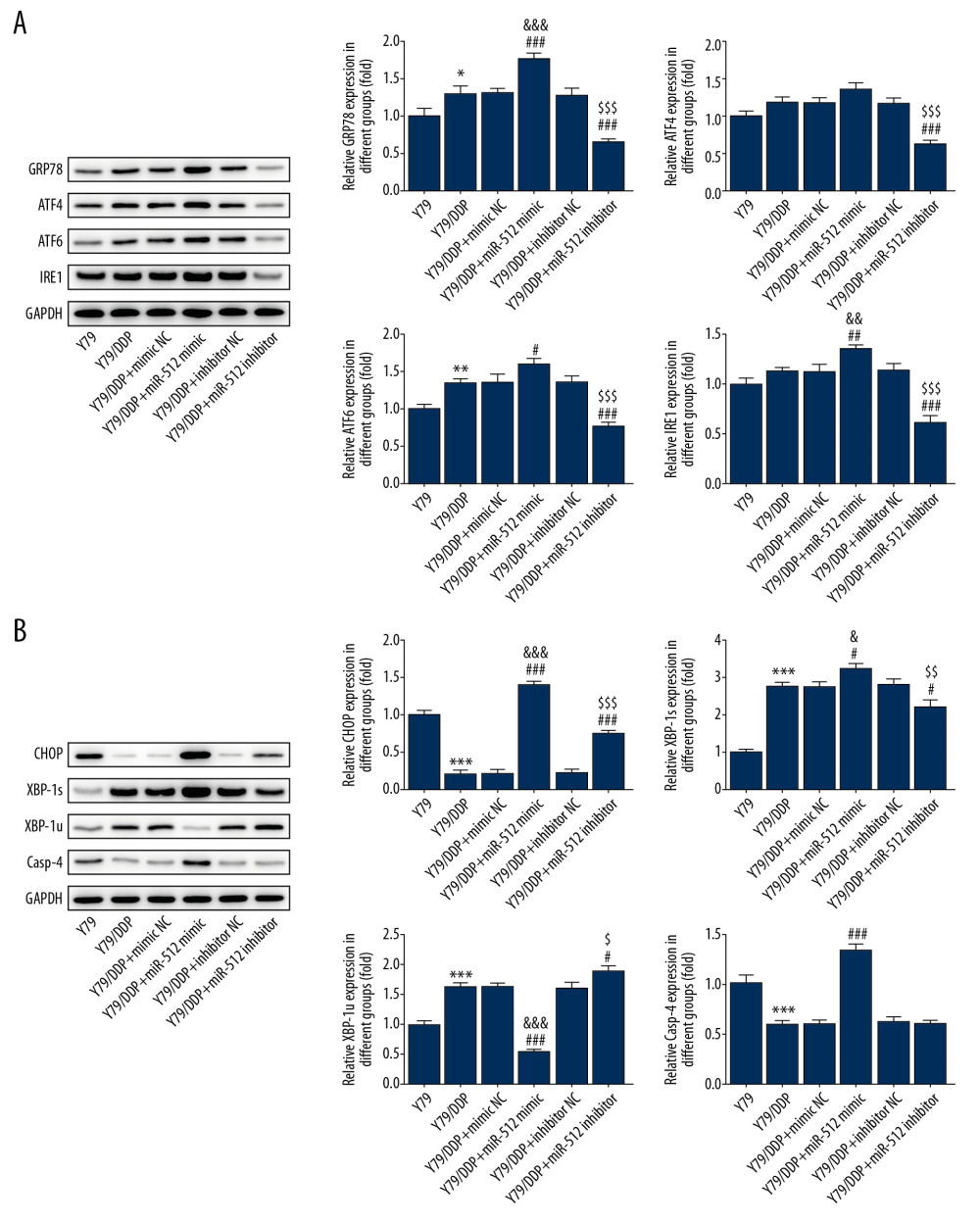 Figure 5. miR-512-3p affects the related proteins expression in ERS. (A) The expression of GRP78, ATF4, ATF6 and IRE1 in Y79/DDP cells after transfection was determined by Western blot analysis. * P<0.05 and ** P<0.05 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. && P<0.01 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of CHOP, XBP-1s, XBP-1u and Casp-4 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 vs. Y79/DDP+inhibitor NC group.
Figure 5. miR-512-3p affects the related proteins expression in ERS. (A) The expression of GRP78, ATF4, ATF6 and IRE1 in Y79/DDP cells after transfection was determined by Western blot analysis. * P<0.05 and ** P<0.05 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. && P<0.01 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of CHOP, XBP-1s, XBP-1u and Casp-4 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 vs. Y79/DDP+inhibitor NC group. 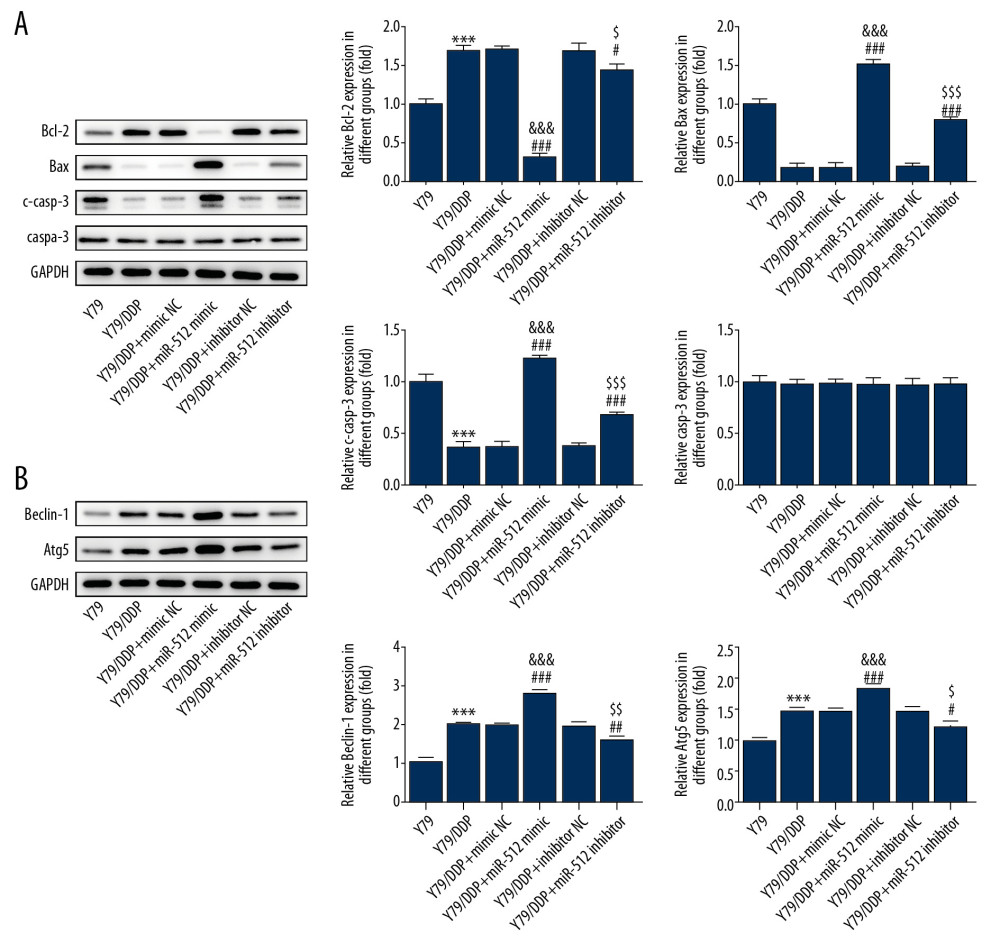 Figure 6. miR-512-3p affects the related proteins expression in apoptosis and autophagy. (A) The expression of casp-3, c-casp-3, Bax and Bcl-2 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of Beclin-1 and Atg5 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$ P<0.01 vs. Y79/DDP+inhibitor NC group.
Figure 6. miR-512-3p affects the related proteins expression in apoptosis and autophagy. (A) The expression of casp-3, c-casp-3, Bax and Bcl-2 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of Beclin-1 and Atg5 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$ P<0.01 vs. Y79/DDP+inhibitor NC group. 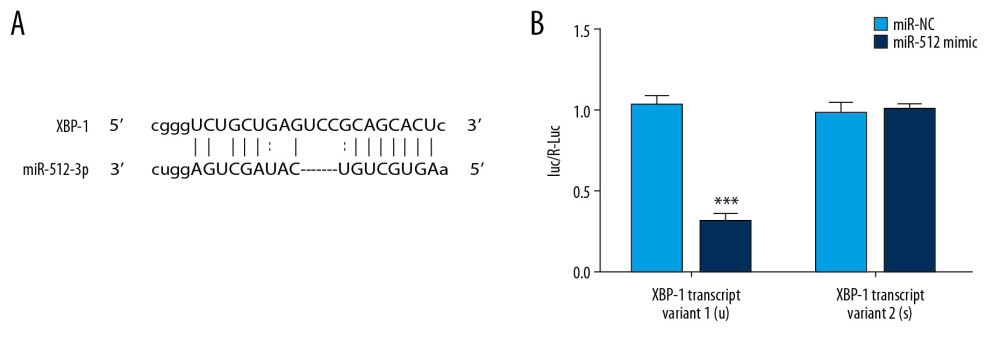 Figure 7. miR-512-3p is combined with XBP-1 transcript variant 1. (A) The binding sites of miR-512-3p and XBP-1 were predicted by ENCORI. (B) Luciferase activity was analyzed in cells co-transfected with miR-512-3p mimic or miR-NC and XBP-1 transcript variant 1 or XBP-1 transcript variant 2. *** P<0.001 vs. miR-NC group.
Figure 7. miR-512-3p is combined with XBP-1 transcript variant 1. (A) The binding sites of miR-512-3p and XBP-1 were predicted by ENCORI. (B) Luciferase activity was analyzed in cells co-transfected with miR-512-3p mimic or miR-NC and XBP-1 transcript variant 1 or XBP-1 transcript variant 2. *** P<0.001 vs. miR-NC group. References
1. Kivelä T, The epidemiological challenge of the most frequent eye cancer: Retinoblastoma, an issue of birth and death: Br J Ophthalmol, 2009; 93; 1129-31
2. Zhao J, Li S, Shi J, Wang N, Clinical presentation and group classification of newly diagnosed intraocular retinoblastoma in China: Br J Ophthalmol, 2011; 95; 1372-75
3. Lin P, O’Brien J, Frontiers in the management of retinoblastoma: Am J Ophthalmol, 2009; 148; 192-98
4. Chintagumpala M, Chévez-Barrios P, Paysse E, Review of current management: Oncologist, 2007; 12; 1237-46
5. Kong X, Modern treatment of retinoblastoma: Ophthalmology in China, 2004; 13; 325-30
6. Shields CL, Sun H, Zhang JProgress in the treatment of retinoblastoma: Chinese Journal of Ocular Fundus Diseases, 2004; 020; 194-97 [in Chinese]
7. Yadar Rk, Chae S-W, Kim HR, Chae H, Endoplasmic reticulum stress and cancer: J Cancer Prev, 2014; 19; 75-88
8. Martinon F, Targeting endoplasmic reticulum signaling pathways in cancer: Acta Oncol, 2012; 51; 822-30
9. Travers KJ, Patil CK, Wodicka L, Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation: Cell, 2000; 101; 249-58
10. Tsai YC, Weissman A, The unfolded protein response, degradation from endoplasmic reticulum and cancer: Genes Cancer, 2010; 1; 764-78
11. Wang M, Kaufman R, Protein misfolding in the endoplasmic reticulum as a conduit to human disease: Nature, 2016; 529; 326-35
12. Ma J, Wang J, Zhang S, The unfolded protein response and diabetic retinopathy: J Diabetes Res, 2014; 2014 160140
13. Eizirik D, Miani M, Cardozo A, Signalling danger: Endoplasmic reticulum stress and the unfolded protein response in pancreatic islet inflammation: Diabetologia, 2012; 56(2); 234-41
14. Shen X, Ellis RE, Lee K: Cell, 2001; 107; 893-903
15. Calfon M, Zeng H, Urano F, IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA: Nature, 2002; 415; 92-96
16. Yoshida H, Matsui T, Yamamoto A, XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor: Cell, 2001; 107; 881-91
17. Dewal M, DiChiara A, Antonopoulos A, XBP1s links the unfolded protein response to the molecular architecture of mature N-glycans: Chem Biol, 2015; 22; 1301-12
18. Chae U, Park S-J, Kim B, Critical role of XBP1 in cancer signaling is regulated by PIN1: Biochem J, 2016; 473(17); 2603-10
19. Jin C, Jin Z, Chen N-Z, Activation of IRE1 alpha-XBP1 pathway induces cell proliferation and invasion in colorectal carcinoma: Biochem Biophys Res Commun, 2016; 470; 75-81
20. Liu Y, Hou X, Liu M, XBP1 silencing decreases glioma cell viability and glycolysis possibly by inhibiting HK2 expression: J Neurooncol, 2015; 126(3); 455-62
21. Wang K, Sun H, Sun T, Long non-coding RNA AFAP1-AS1 promotes proliferation and invasion in prostate cancer via targeting miR-512-3p: Gene, 2020; 726; 144169
22. Zhu X, Gao G, Chu K, Inhibition of RAC1-GEF DOCK3 by miR-512-3p contributes to suppression of metastasis in non-small cell lung cancer: Int J Biochem Cell Biol, 2015; 61; 103-14
23. Chen F, Zhu H-H, Zhou L-F, Inhibition of c-FLIP expression by miR-512-3p contributes to taxol-induced apoptosis in hepatocellular carcinoma cells: Oncol Rep, 2010; 23; 1457-62
24. Rowland AA, Voeltz GK, Endoplasmic reticulum-mitochondria contacts: Function of the junction: Nat Rev Mol Cell Biol, 2012; 13; 607-25
25. Marchi S, Patergnani S, Pinton P, The endoplasmic reticulum-mitochondria connection: One touch, multiple functions: Biochim Biophys Acta, 2013; 1837(4); 461-69
26. Berridge M, The endoplasmic reticulum: A multifunctional signaling organelle: Cell Calcium, 2002; 32; 235-49
27. Schonthal A, Endoplasmic reticulum stress: Its role in disease and novel prospects for therapy: Scientifica, 2012; 2012 857516
28. Rademakers S, Span P, Kaanders J, Molecular aspects of tumour hypoxia: Mol Oncol, 2008; 2; 41-53
29. Li L, Li G, Wei H, The endoplasmic reticulum stress response is associated with insulin resistance-mediated drug resistance in HepG2 cells: Neoplasma, 2015; 62; 180-90
30. Fu Y-F, Liu X, Gao M, Endoplasmic reticulum stress induces autophagy and apoptosis while inhibiting proliferation and drug resistance in multiple myeloma through the PI3K/Akt/mTOR signaling pathway: Oncotarget, 2017; 8(37); 61093-106
31. Nikesitch N, Lee J, Ling S, Roberts T, Endoplasmic reticulum stress in the development of multiple myeloma and drug resistance: Clin Transl Immunol 7, 2018; 7(1); e1007
32. Shi Z, Yu X, Yuan M, Activation of the PERK-ATF4 pathway promotes chemo-resistance in colon cancer cells: Sci Rep, 2019; 9(1); 3210
33. Xiu W, Yi H, Guodong H, Endoplasmic reticulum stress induces miR-706, a pro-cell death microRNA, in a protein kinase RNA-Like ER kinase (PERK) and activating transcription factor 4 (ATF4) dependent manner: Cell J, 2020; 22; 394-400
34. Luhr M, Torgersen M, Lindner P, The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress: J Biol Chem, 2019; 294(20); 8197-217
35. Zinszner H, Kuroda M, Wang X, CHOP is implicated in programmed cell death in response to impaired function of the endoplasmic reticulum: Genes Dev, 1998; 12; 982-95
36. Ding W-X, Ni H-M, Gao W, Differential effects of endoplasmic reticulum stress-induced autophagy on cell survival: J Biol Chem, 2007; 282; 4702-10
37. Rouschop KMA, van den Beucken T, Dubois L, The unfolded protein response protects human tumor cells during hypoxia through regulation of the autophagy genes MAP1LC3B and ATG5: J Clin Invest, 2010; 120; 127-41
38. Ogata M, Hino S-i, Saito A, Autophagy is activated for cell survival after endoplasmic reticulum stress: Mol Cell Biol, 2006; 26; 9220-31
39. Chen R, Dai R, Duan C, Unfolded protein response suppresses cisplatin-induced apoptosis via autophagy regulation in human hepatocellular carcinoma cells: Folia Biol (Praha), 2011; 57; 87-95
Figures
Figure 1. DDP-resistant cells were constructed by gradient concentration of DDP. (A) The DDP inhibition rates for DDP-resistant RB cells were reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group. (B) The expression of XBP-1 in DDP-resistant RB cells was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.001 vs. weri-RB1 group. &&& P<0.001 vs. HXO-RB44 group.Figure 2. DDP-resistant Y79 cells (Y79/DDP cells) were transfected. RT-PCR analysis confirmed the transfection effects. *** P<0.001 vs. control group. ### P<0.001 vs. mimic NC group. &&& P<0.01 vs. inhibitor NC group.Figure 3. Proliferation of Y79/DDP cells and DDP inhibition rate for Y79/DDP cells were changed after transfection. (A) The proliferation of Y79/DDP cells after transfection was detected by CCK-8 assay. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group. (B) The DDP inhibition rates for Y79/DDP cells after transfection were also reflected by CCK-8 assay. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group.Figure 4. miR-512-3p affects the apoptosis and autophagy of Y79/DDP cells. (A) The apoptosis of Y79/DDP cells after transfection was determined by TUNEL assay. (B) The proteins expression of autophagy in Y79/DDP cells after transfection was detected by Western blot analysis. *** P<0.001 vs. Y79 group. ## P<0.01 and ### P<0.001 vs. Y79/DDP group. ΔΔΔ P<0.001 vs. Y79/DDP+mimic NC group. $$ P<0.01 vs. Y79/DDP+inhibitor NC group.Figure 5. miR-512-3p affects the related proteins expression in ERS. (A) The expression of GRP78, ATF4, ATF6 and IRE1 in Y79/DDP cells after transfection was determined by Western blot analysis. * P<0.05 and ** P<0.05 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. && P<0.01 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of CHOP, XBP-1s, XBP-1u and Casp-4 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 vs. Y79/DDP+inhibitor NC group.Figure 6. miR-512-3p affects the related proteins expression in apoptosis and autophagy. (A) The expression of casp-3, c-casp-3, Bax and Bcl-2 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. ### P<0.001 vs. Y79/DDP group. &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$$ P<0.001 vs. Y79/DDP+inhibitor NC group. (B) The expression of Beclin-1 and Atg5 in Y79/DDP cells after transfection was determined by Western blot analysis. *** P<0.001 vs. Y79 group. # P<0.05, ## P<0.01 and ### P<0.001 vs. Y79/DDP group. & P<0.05 and &&& P<0.001 vs. Y79/DDP+mimic NC group. $ P<0.05 and $$ P<0.01 vs. Y79/DDP+inhibitor NC group.Figure 7. miR-512-3p is combined with XBP-1 transcript variant 1. (A) The binding sites of miR-512-3p and XBP-1 were predicted by ENCORI. (B) Luciferase activity was analyzed in cells co-transfected with miR-512-3p mimic or miR-NC and XBP-1 transcript variant 1 or XBP-1 transcript variant 2. *** P<0.001 vs. miR-NC group. In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387