24 November 2020: Animal Study
Exploring the Mechanism of Icariin in Osteoporosis Based on a Network Pharmacology Strategy
Zhiyong Long12ABCDE, Jiamin Wu3ABCDE, Wang Xiang4ABCDE, Zhican Zeng5ABCDE, Ganpeng Yu6ABCDE*, Jun Li6ABCDDOI: 10.12659/MSM.924699
Med Sci Monit 2020; 26:e924699






Abstract
BACKGROUND: With the aging of the world’s population, the incidence of osteoporosis (OP) has become a public health problem of worldwide concern. Research shows that icariin may have a therapeutic effect on OP.
MATERIAL AND METHODS: PharmMapper was utilized to predict the potential targets of icariin. GeneCards and Online Mendelian Inheritance in Man (OMIM) were used for the collection of OP genes. The STRING database was utilized to obtain the protein-protein interaction (PPI) data. We used Cytoscape 3.7.2 to construct and analyze the networks. The genes and targets in the networks were input into the Database for Annotation, Visualization and Integrated Discovery (DAVID) to undergo Gene Ontology (GO) and pathway enrichment analysis. Finally, animal experiments were performed to verify the prediction results of this study.
RESULTS: A total of 297 icariin potential targets and 262 OP genes were obtained, and an icariin-OP PPI network was constructed and analyzed. The results of the GO enrichment analysis showed that icariin can regulate the steroid hormone-mediated signaling pathway, skeletal system development, extracellular space, cytosol, and steroid hormone receptor activity. The results of the pathway enrichment analysis showed that icariin can regulate osteoclast differentiation, FoxO, estrogen, and PPAR signaling pathways. The results of the experiments showed that icariin can increase estradiol, β-catenin, and Receptor Activator of Nuclear Factor-к B Ligand (RANKL)/osteoprotegerin (OPG) ratio in postmenopausal OP rats (P<0.05).
CONCLUSIONS: This research found that the icariin can regulate OP-related biological processes, cell components, molecular functions, and signaling pathways.
Keywords: Osteoporosis, Osteoporosis, Postmenopausal, Pharmacology, phytochemicals, Bone Density, Femur, gene ontology, Protein Interaction Maps, RANK Ligand, beta Catenin
Background
With the aging of the world’s population, the incidence of osteoporosis (OP) has been increasing year by year, and it has become a public health problem of worldwide concern [1,2]. Primary OP is divided into type I and type II; type I OP is postmenopausal osteoporosis (PMOP) and type II OP is senile osteoporosis [3]. PMOP is caused by endocrine disorders due to hypoovarian function after menopause, especially the decrease in estrogen level. Because bone resorption is greater than bone formation, osteoblasts required for bone reconstruction are relatively reduced. The relative increase in osteoclasts leads to a decrease in systemic bone mass and destruction of bone tissue microstructure, resulting in increased bone fragility and metabolic bone disease, predisposing bones to fracture [3–5]. At present, the drugs recognized for the treatment of OP mainly include bone resorption inhibitors, bone formation promoters, and bone minerals [6]. However, from the perspective of drug treatment effects, the current drugs only improve clinical symptoms and delay the progression of the disease, and they cannot restore the patient’s bone remodeling process to a balanced state. Therefore, it is urgent to find new ideas to solve this problem [7,8].
Traditional Chinese medicine (TCM) has a role in treating OP [9]. Modern pharmacological studies have found that herbal formulas for treating OP have the combined effects of improving intestinal calcium absorption, promoting osteoblast mineralization, enhancing cell osteogenic activity, and inhibiting the formation of osteoclasts [9,10]. Hence, the active ingredients of herbal medicines have unique advantages and broad development prospects in the prevention and treatment of OP. Icariin is a compound derived from
Material and Methods
POTENTIAL TARGETS PREDICTION OF ICARIIN AND OP GENES COLLECTION:
The molecular structure of icariin was queried at PubChem (https://pubchem.ncbi.nlm.nih.gov/), and its SDF file can be downloaded from this website. Then, the SDF file was input into the PharmMapper server (http://lilab-ecust.cn/pharmmapper/) to predict the potential targets of icariin [16]. Finally, a total of 297 of icariin’s potential targets were predicted by PharmMapper (Supplementary Table 1, see supplementary materials).
The GeneCards (http://www.genecards.org) and Online Mendelian Inheritance in Man (OMIM) (http://omim.org/) databases were utilized to collect the OP genes [17,18]. After the search, a total of 2983 OP-related genes were obtained. The genes with relevance score ≥8.0 were selected for sequence research (Supplementary Table 2).
NETWORK CONSTRUCTION AND ANALYSIS METHODS:
The protein-protein interaction (PPI) data of icariin targets and OP genes were collected from STRING 11.0 (http://string-db.org/) [19]. After entering the website, select “Multiple Proteins”, we imported icariin targets and OP genes into “List of Names”, set Organism to “Homo sapiens” for PPI, and then downloaded the file in “tsv” format. Cytoscape 3.7.2 software (https://cytoscape.org/) was utilized to draw the networks, such as icariin-OP PPI network, and to perform network analysis [20].
The targets and genes in the icariin-OP PPI network were input into the Database for Annotation, Visualization and Integrated Discovery (DAVID) 6.8 (https://david.ncifcrf.gov/) to undergo Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis [21].
ANIMALS:
Forty-eight (48) specific pathogen-free (SPF)-grade female SD rats, weighing 220–250 g, 10–12 months old, were purchased from Hunan Slake Jingda Experimental Animal Co., Ltd. [Certificate No. 43004700005517, License No. SCXK (Xiang) 2013–0004]. The animals were housed in an air-conditioned room with constant temperature (23±2°C) and humidity (45–50%), and a 12h/12h light/dark cycle. These animals were housed at the Experimental Animal Management Center of People’s Hospital of Ningxiang (Ningxiang, China), Hunan University of Chinese Medicine. This study was approved by the Ethics Committee of People’s Hospital of Ningxiang (Ningxiang, China), Hunan University of Chinese Medicine (HUCM-0018). All animal care and experimental procedures were in accordance with the National Institute of Health’s Guide for the Care and Use of Laboratory Animals.
INSTRUMENTS AND REAGENTS:
We used the following instruments and reagents: Estradiol (E2) enzyme-linked immunosorbent assay (ELISA) Kit (Shanghai Bangyi Biotechnology Co., Ltd.), rat osteoprotectin (OPG) ELISA test kit, rat β-catenin ELISA kit, rat nuclear factor κB receptor activating factor ligand (RANKL) ELISA test kit (Suzhou Calvin Biotechnology Co., Ltd), TRIZOL reagent (Invitrogen, Inc.), Reverse Transcription System, Go Taq Green Master Mix (Promega, Inc.), liquid nitrogen (Changsha Rizhen Gas Co., Ltd.), Estradiol valerate (Bayer Healthcare Co., Ltd. Guangzhou Branch), chloroform, isopropanol (Sinopharm Group Chemical Reagent Co., Ltd.), icariin (content greater than 98%) (Shanghe Biotechnology Co., Ltd.), Eppendorf BioPhotometer Plus Nucleic Acid Protein Analyzer (Eppendorf Inc.); InfiniteF50 microplate reader (Tecan Inc.), gel imaging analysis system (Bio-Rad, Inc.), Horizontal electrophoresis apparatus (Beijing Liuyi Biotechnology Co., Ltd.), Electronic balance (Ohaus Instrument Co., Ltd.), Enzyme-labeled instrument (MB53Q) (Shenzhen Huisong Technology Development Co., Ltd.), and low-temperature high-speed centrifuge (1–14VK) (Sigma Inc.).
ANIMAL MODELING, GROUPING, AND INTERVENTION:
The PMOP pathological model of adult female rats with ovariectomy for 3 months was used in this study.
MODELING METHOD:
Rats were anesthetized with 2% sodium pentobarbital (0.2 mL/100 g body weight), and under sterile conditions, the skin and both sides of the muscles were longitudinally excised at a distance of 1 cm from the outside of the thoracolumbar vertebrae of the rats, and bilateral ovaries were removed. The wound was sutured in 2 layers and the blood stains were scrubbed with saline. Penicillin sodium (40 000 units each) was intramuscularly injected for 3 consecutive days after the operation, and the sutures were removed 5 days after the operation. The animals were kept in a clean environment at room temperature of 23–25°C and relative humidity of 40~60%. In the sham operation group, we only removed the corresponding volume of fat around the ovary.
ANIMAL GROUPING:
The rats were randomly divided into 4 groups: the sham operation group, the model group, the icariin group [150 mg/kg/d)], and the E2 group [0.167 mg/kg/d] according to body weight, with 12 rats in each group. All groups except the sham operation group were resected with bilateral ovaries to construct PMOP pathological models. Interventions started 1 week after surgery.
INTERVENTION METHODS:
Rats in the icariin group were orally administered icariin 150 mg/kg daily, and rats in the E2 group were orally administered E2 0.167 mg/kg daily. Rats in the sham operation group and the model group were intragastrically administered an equal volume of 0.9% sodium chloride daily. The intervention lasted 12 weeks.
SPECIMEN COLLECTION:
After the intervention, 8 female rats were randomly selected from each group for testing. All rats were anesthetized by intraperitoneal injection of 2% pentobarbital (0.2 mL/100g body weight), and blood was collected through the abdominal aorta. The blood sample was naturally coagulated at room temperature, and then the serum was separated by centrifugation (1000 g), and stored at −80°C for later use. The left femur of the rat was stripped, the muscles on the bone were removed, and the periosteum was retained for pathomorphological observation.
BONE MINERAL DENSITY (BMD) MEASUREMENT:
After taking blood from the abdominal aorta, the 2nd to 4th lumbar vertebrae of the rats were collected and the muscles on the bones were removed; the samples were wrapped with 0.9% sodium chloride gauze without paraformaldehyde and stored at −4°C. The samples were later sent to the Bone Density Test Room of People’s Hospital of Ningxiang, Hunan University of Chinese Medicine to measure the BMD of the lumbar spine.
DETERMINATION OF SERUM E2, β-CATENIN, RANKL, AND OPG LEVELS:
The serum E2, β-catenin, RANKL, and OPG levels were detected by enzyme-linked immunosorbent assay (ELISA). The serum samples were coagulated at room temperature for 20 min and centrifuged at 3000 rpm for 10 min. The supernatant was collected and stored at −20°C. Then, the standard was diluted and loaded. After that, blank wells and experimental wells were set, in which 40 μL of sample dilution and 10 μL of samples were added to the experimental wells. Subsequently, the enzyme-labeled coated plate was incubated at 37°C for 30 min. After washing the plate, 50 μL of enzyme-labeled reagent was added to each well (except the blank well) and incubated at 37°C for 30 min. After washing the plate again, 50 μL of A and B color reagents were added to each well and incubated at 37°C in the dark for 15 min. Finally, a stop solution was added to each well to stop the reaction, and the optical density (OD) value was measured at 450 nm.
STATISTICAL ANALYSIS:
Statistical analysis was performed using SPSS 22.0 software. The experimental data were expressed as mean±SD (x±SD) and the data that conformed to normal distribution were analyzed by one-way ANOVA (for comparison of differences between groups) or
Results
ICARIIN POTENTIAL TARGETS AND OP GENES:
A total of 297 icariin potential targets and 262 OP genes were obtained. There are 22 identical targets between the 2 target sets (Figure 2). The PPI data of icariin targets and OP genes were collected from STRING 11.0 for sequence research.
ICARIIN-OP PPI NETWORK:
The icariin targets, OP genes, and PPI data were input into Cytoscape 3.7.2 to construct an icariin-OP PPI network. This network is composed of 213 OP gene nodes, 272 icariin target nodes, 22 icariin-OP target nodes and 7823 edges (Figure 3). The nodes were sorted according to their degree, and the top 20 were: (1) Icariin targets: AKT1 (188 edges), EGFR (157 edges), MAPK1 (138 edges), MMP9 (137 edges), MAPK8 (123 edges), CASP3 (119 edges), HSP90AA1 (100 edges); (2) OP genes: INS (235 edges), IL6 (200 edges), TNF (170 edges), MAPK3 (150 edges), CTNNB1 (131 edges), JUN (131 edges), IL1B (117 edges), LEP (110 edges), SPP1 (106 edges); (3) Icariin-OP targets: ALB (227 edges), IGF1 (151 edges), SRC (136 edges), and ESR1 (129 edges).
GO ENRICHMENT ANALYSIS RESULTS OF ICARIIN-OP PPI NETWORK:
The targets and genes in icariin-OP PPI network were input into DAVID to undergo GO enrichment analysis and returned many biological processes, cell components, and molecular functions (Figure 4). The results of the GO enrichment analysis were further screened to obtain OP-related biological processes, cell components, and molecular functions. They were sorted according to their degree of enrichment (negative correlation with P value) and counts.
The top 10 OP-related biological processes were: steroid hormone-mediated signaling pathway, skeletal system development, positive regulation of cell proliferation, negative regulation of apoptotic process, ossification, extracellular matrix disassembly, collagen catabolic process, peptidyl-tyrosine autophosphorylation, response to glucocorticoid, and bone resorption (Figure 5, Supplementary Table 3)
The top 10 OP-related cell components were: extracellular space, extracellular region, extracellular exosome, cytosol, extracellular matrix, proteinaceous extracellular matrix, extrinsic component of the cytoplasmic side of plasma membrane, receptor complex, membrane raft, and endoplasmic reticulum lumen (Figure 6, Supplementary Table 4).
The top 10 OP-related molecular function were: steroid hormone receptor activity, RNA polymerase II transcription factor activity, ligand-activated sequence-specific DNA binding, protein homodimerization activity, growth factor activity, identical protein binding, serine-type endopeptidase activity, protein tyrosine kinase activity, receptor binding, cytokine activity, and steroid binding (Figure 7, Supplementary Table 5).
KEGG PATHWAYS ENRICHMENT ANALYSIS RESULTS OF ICARIIN-OP PPI NETWORK:
The targets and genes in the icariin-OP PPI network were input into DAVID to undergo KEGG pathway enrichment analysis and returned many signaling pathways (Figure 8). The results of KEGG pathway enrichment analysis were further screened to obtain OP-related signaling pathways. They were sorted according to their degree of enrichment (negative correlation with P value) and counts. The top 10 signaling pathways were: Osteoclast differentiation, FoxO signaling pathway, Estrogen signaling pathway, PPAR signaling pathway, Ovarian steroidogenesis, PI3K-Akt signaling pathway, T cell receptor signaling pathway, TNF signaling pathway, Insulin resistance, and MAPK signaling pathway (Figure 9, Supplementary Table 6). The role of each target in the Wnt and NF-κB signaling pathway is shown in Figure 10.
Based on the prediction results of network pharmacology, we found that icariin can regulate multiple signaling pathways through core targets, thereby directly and indirectly regulating bone metabolism to exert an anti-OP effect. Next, the prediction results were further verified through animal experiments.
EFFECT OF ICARIIN ON BMD:
The difference in lumbar BMD between the sham operation group and the model group was statistically significant (P<0.05), indicating that the animal pathological model of PMOP was successful. Compared with the model group, the BMD of the icariin group increased (P<0.05), but compared with the estrogen group, there was no statistically significant difference (P> 0.05) (Figure 11).
EFFECT OF ICARIIN ON MORPHOLOGY OF LEFT FEMUR BONE IN RATS:
In the sham operation group, the trabecular bones were evenly distributed, and the tibial trabecular bone density and trabecular bone area/medullary cavity area ratio were normal. In the model group, the trabecular bone was sparse and damaged, the bone marrow cavity was enlarged, and the trabecular bone density and the trabecular bone area/medullary cavity area ratio were significantly decreased. In the icariin group, the trabecular bone distribution was relatively uniform, and the trabecular bone density was roughly restored (Figure 12).
EFFECT OF ICARIIN ON SERUM E2 AND β-CATENIN LEVEL AND RANKL/OPG RATIO:
Compared with the model group, the serum E2 levels of the rats in the sham operation group, icariin group, and E2 group were all increased, and the differences were statistically significant (
Compared with the sham operation group, the serum β-catenin level in the model group was significantly decreased (P<0.01). Compared with the model group, the serum β-catenin level in the E2 group was significantly increased (P<0.01), and the serum β-catenin level in the icariin group was also increased (P<0.05) (Figures 13, 14).
Compared with the sham operation group, the serum RANKL/OPG ratio in the model group was significantly increased (P<0.01). Compared with the model group, the serum RANKL/OPG ratio of the icariin group and the E2 group was significantly decreased (P<0.01) (Figures 15, 16).
Discussion
This research constructed and analyzed the icariin-OP PPI network. The results showed that the icariin potential targets can interact with many OP genes in the OP disease target network. Targets such as ALB, IGF1, SRC, and ESR1 are at the center of the network; these targets play a crucial role in the OP’s pathological process. For example, IGF-1 plays a key role in the balance between bone resorption and bone formation. During bone resorption, osteoclasts produce IGF-1 and stimulate new bone formation [22]. In addition, IGF-1 also inhibits osteoblast apoptosis [23]. During bone formation, IGF-1 and IGFBP synthesis and secretion increase. IGF-1 binds to osteoblast surface receptors to increase collagen synthesis and inhibit collagenase production in osteoclasts. IGF-1 can also acidify osteoblasts and bone matrix, which is beneficial to bone mineralization [24,25]. ESR1, a type of ligand-activated nuclear transcription factor, mediates most of the estrogen response and is a nuclear macromolecule that mediates the biological role of estrogen [26]. Estrogen is an important factor in increasing bone density and preventing bone loss after menopause. ESR was found on the surface of both human osteoblasts and osteoclasts, which indicates that estrogen plays a direct regulatory role on bone cells [27,28].
From the results of GO enrichment, we found that icariin mainly regulates the production of estrogen, steroid hormones and their signaling pathways, bone development, proliferation and apoptosis, ossification, and bone resorption. The above biological processes play an important role in the physiological processes of ossification and bone resorption. The core signaling pathways are FoxO, estrogen, PPAR, PI3K-Akt, Wnt, NF-κB, T cell receptor, TNF signaling pathway, and ovarian steroid production. These pathways directly and indirectly affect bone metabolism. The FoxO signaling pathway mainly regulates oxidative stress during osteoporosis. Inhibiting the transcription of FoxO activated by oxidative stress through anti-oxidative stress and positively regulating the Wnt signaling pathway can improve the osteogenic differentiation and bone formation of bone tissue [29,30]. It can also reduce high conversion levels of bone metabolism and reduce bone resorption [29,30]. Estrogen is a steroid hormone produced by the endocrine system. It is involved in the physiological and pathological processes of human bones and plays this important role in bone reconstruction [31]. The estrogen drugs currently used for OP include exogenous estrogen, phytoestrogens (soy isoflavones), and selective estrogen receptor modulators [32,33]. Recent research indicates that the PI3K/Akt signaling pathway inhibits the progression of OP by promoting osteoblast proliferation, differentiation, and bone formation [34]. Wu et al. found that the mechanism of Si-Wu-Tang, a TCM formula used in treating OP, is to activate the PI3K/Akt and NF-κB signaling pathways, increases the expression of ALP, BMP-2, and OPN genes, promote osteoblast differentiation, and increases bone formation [35]. In addition, PI3K and downstream signals RANKL, c-Fms, and αvβ3 integrin play important roles in the processes of osteoclast survival, differentiation, and bone resorption [36]. The Wnt/β-catenin signaling pathway, as an important regulator of bone health and bone disease, can change bone mass through multiple pathways and is closely involved in OP [37,38]. The RANKL/RANK/OPG/NF-κB signaling pathway mainly regulates the differentiation and absorption activity of osteoclasts and participates in physiological and pathological bone reconstruction [39]. RANKL binds to the Receptor Activator of Nuclear Factor-κ B (RANK) receptor on the surface of osteoclasts, allowing TRAF-6 to aggregate in osteoclasts and OCPs, activating IKK/NF-κB, JNK, AP-1, c-Myc, c-Fos, and NFATc1, thereby promoting the differentiation and function of osteoclasts [39,40]. In addition, TGF-beta, BMP, Notch, Hedgehog, MAPK, and other signaling pathways also play important regulatory roles in the occurrence and development of OP [41–43]. These signaling pathways are intertwined with each other, and together influence the occurrence and development of OP.
Icariin, a flavonoid, is an effective pharmacological component extracted from
The classic signaling pathway Wnt/β-catenin also plays an important role in bone. It increases bone mass in a variety of ways, including renewing stem cells, stimulating pre-osteoblastic replication, inducing osteoblastogenesis, and inhibiting osteoblasts and osteoblast apoptosis [52,53]. The key molecule of the Wnt/β-catenin signaling pathway is β-catenin, which is closely involved in the development of bones and is a molecule necessary for osteoblasts to complete the differentiation process [53]. The present study showed that, compared with the sham operation group, the serum β-catenin level in the model group was significantly lower. After treatment with icariin, rat serum β-catenin levels were higher than in the model group, suggesting that icariin activates the Wnt/β-catenin signaling pathway through this effect and plays a role in treating PMOP.
The RANKL/RANK/OPG system is an important signaling pathway that was recently discovered; it plays a role in bone reconstruction and osteoclast differentiation [54]. RANK is an agonist that can bind to RANKL on the surface of osteoclast precursor cells. When RANK is activated, osteoclasts begin to differentiate and mature, and an abundance of activated osteoclasts causes osteolytic bone resorption [55]. OPG is a soluble inhibitory protein and a competitive inhibitor of RANKL; it inhibits the differentiation and maturation of osteoclast precursor cells by competing with RANKL. The expression levels of RANKL and OPG reflect the degree of bone resorption [56]. Generally speaking, when the ratio of RANKL/OPG increases, the number and activity of osteoclasts will increase; when the RANKL/OPG ratio decreases, the number and activity of osteoclasts will decrease. The RANKL/OPG ratio is the decisive factor for the final effect of the RANKL/RANK/OPG system [57]. The present study shows that, compared with the sham operation group, the RANKL/OPG ratio of the model group rats increased significantly. After icariin treatment, the RANKL/OPG ratio decreased more obviously, indicating that icariin can affect the RANKL/RANK/OPG system, and thus can play a role in the treatment of PMOP. In addition to RANKL and OPG, there are other signaling molecules related to OP, such as CTX-1, ALP, TRACP 5b, and PINP [58–60]. In the future, our group plans to further study these OP-related signaling molecules and pathways.
The strength of the present study is that it elucidates the regulatory effect of icariin on the OP biomolecular network, and discovered OP-related targets, biological modules (i.e., clusters), and signaling pathways. Among them, most of the biological modules and signal pathways (such as the regulation of osteoblasts and osteoclasts) have been verified by previous studies; therefore, we chose a new direction that has not yet been studied to verify the prediction results, assessing Wnt pathway and RNKL pathway expression in serum. The limitation of this study is that the prediction results were mainly verified by animal experiments, and relevant clinical trials have not yet been carried out, so there is a lack of support from clinical evidence.
We constructed a molecular network model of icariin intervening in OP through network pharmacology strategies, and found that icariin may play a therapeutic role through osteoclast differentiation, FoxO, Wnt, Estrogen, PPAR, PI3K-Akt, NF-κB, and T cell receptor signaling pathway. Then, we selected the RANKL/RANK/OPG system and Wnt signaling pathway for experimental verification. The experiment revealed that icariin can treat PMOP by regulating serum E2 levels, Wnt/β-catenin signaling pathway, and RANKL/RANK/OPG system at the level of bone metabolism. Recent research also shows the expression of these 2 pathways in bone tissue after icariin intervention [61]. This discovery provides a more in-depth interpretation of the pathogenesis of PMOP and provides a new theoretical basis for the clinical application of icariin.
Conclusions
This research constructed and analyzed an icariin-OP PPI network and found that icariin can regulate OP-related biological processes, cell components, molecular functions, and signaling pathways. Experiments also showed that it can affect PMOP rats by regulating estrogen, Wnt/β-catenin, and RANKL/RANK/OPG signaling pathways.
Figures
 Figure 1. The research process.
Figure 1. The research process. 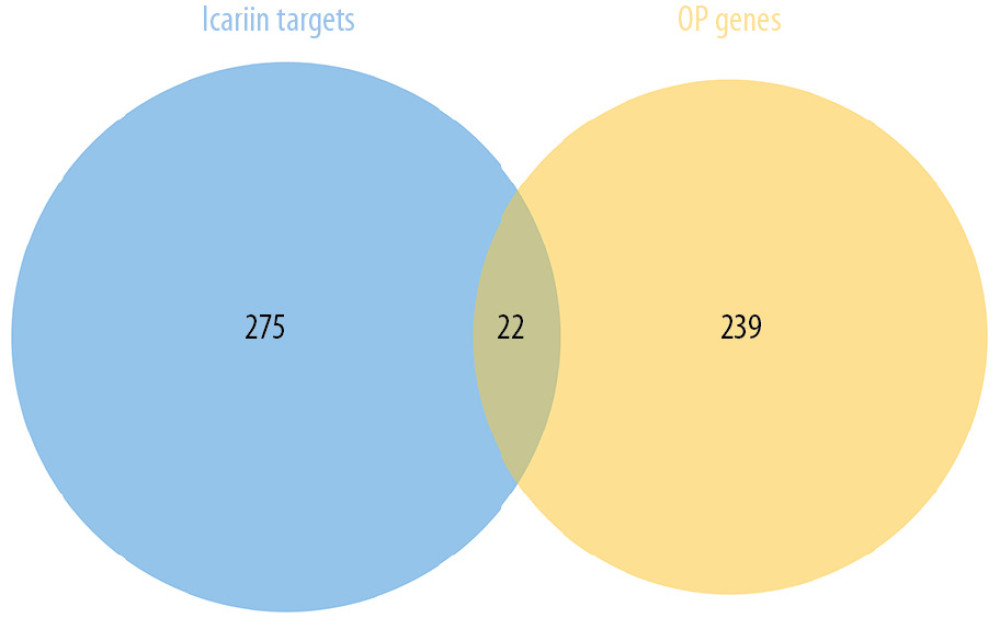 Figure 2. Venn diagram of icariin potential targets and OP genes.
Figure 2. Venn diagram of icariin potential targets and OP genes. 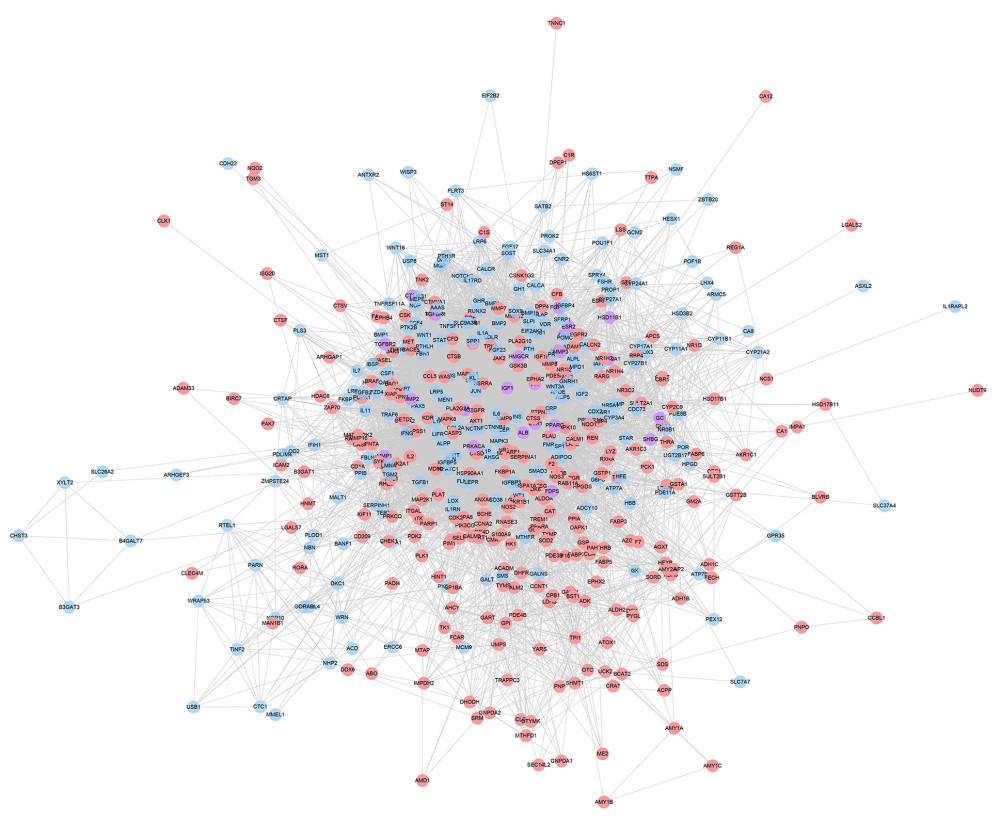 Figure 3. Icariin-OP PPI Network (Pink circles stand for icariin targets. Blue circles stand for OP genes; Purple circles stand for Icariin-OP targets).
Figure 3. Icariin-OP PPI Network (Pink circles stand for icariin targets. Blue circles stand for OP genes; Purple circles stand for Icariin-OP targets). 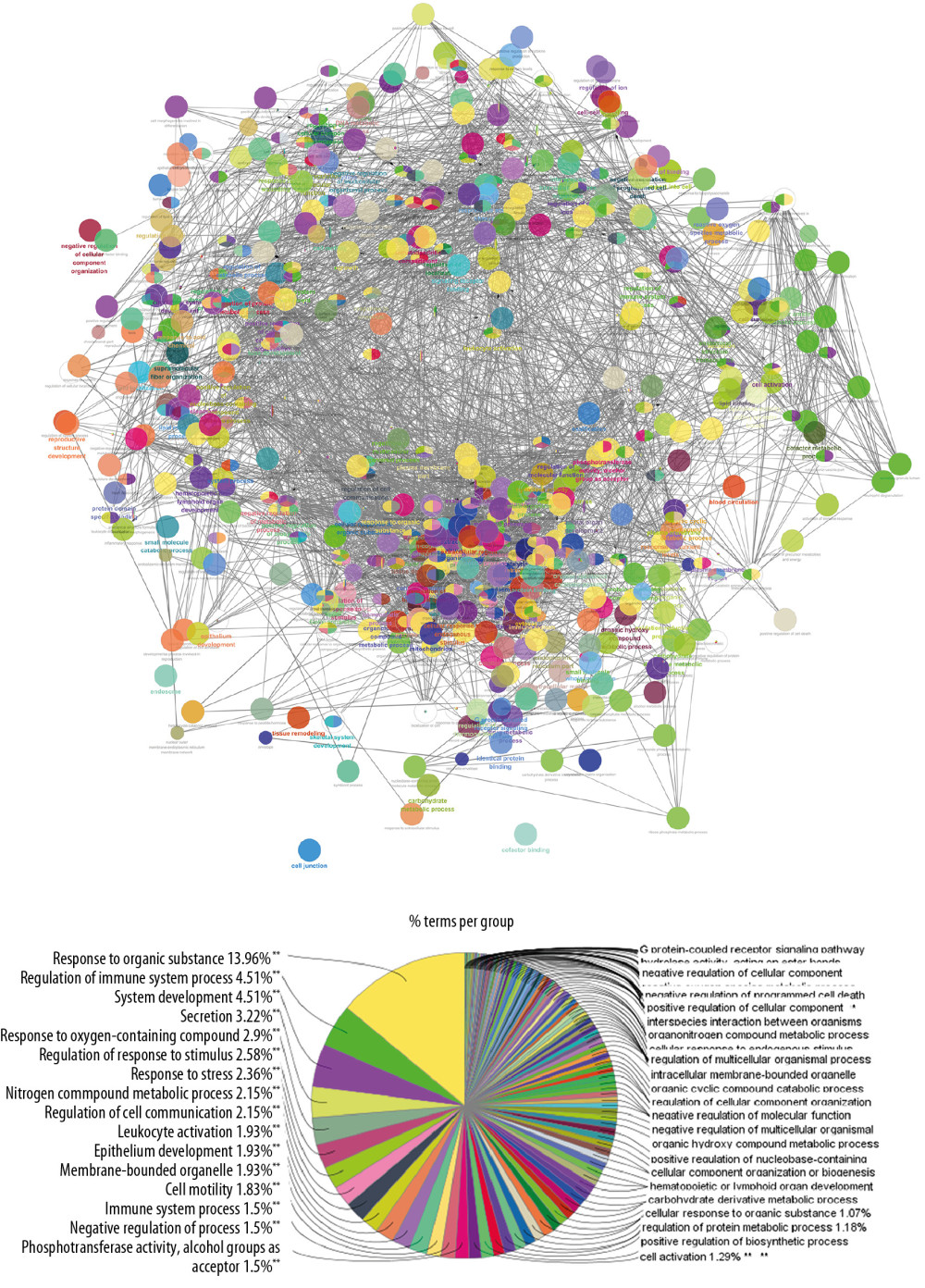 Figure 4. The results of GO enrichment analysis.
Figure 4. The results of GO enrichment analysis. 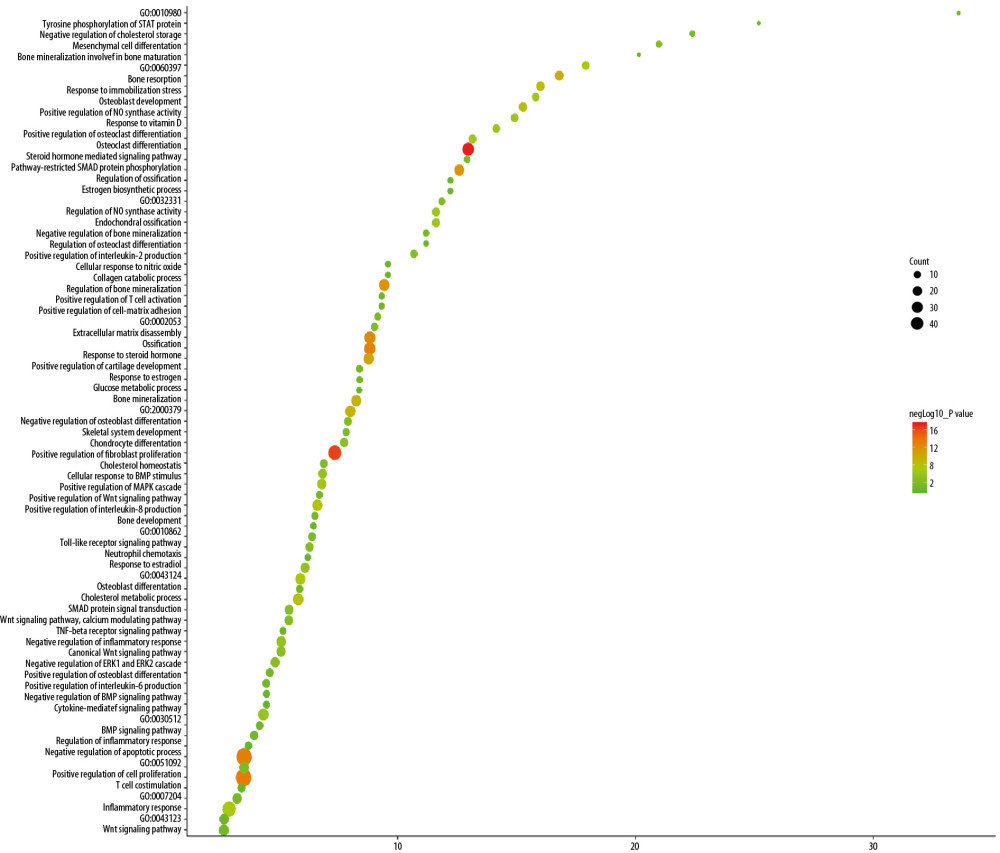 Figure 5. Bubble chart of biological processes (X-axis stand for fold enrichment).
Figure 5. Bubble chart of biological processes (X-axis stand for fold enrichment). 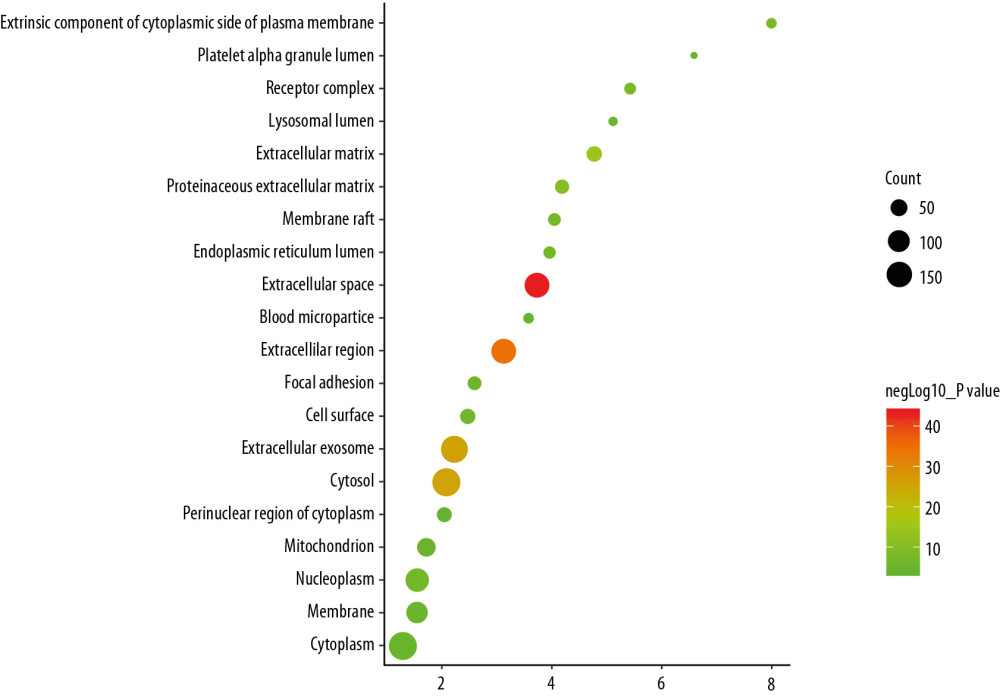 Figure 6. Bubble chart of cell components (X-axis stand for fold enrichment).
Figure 6. Bubble chart of cell components (X-axis stand for fold enrichment). 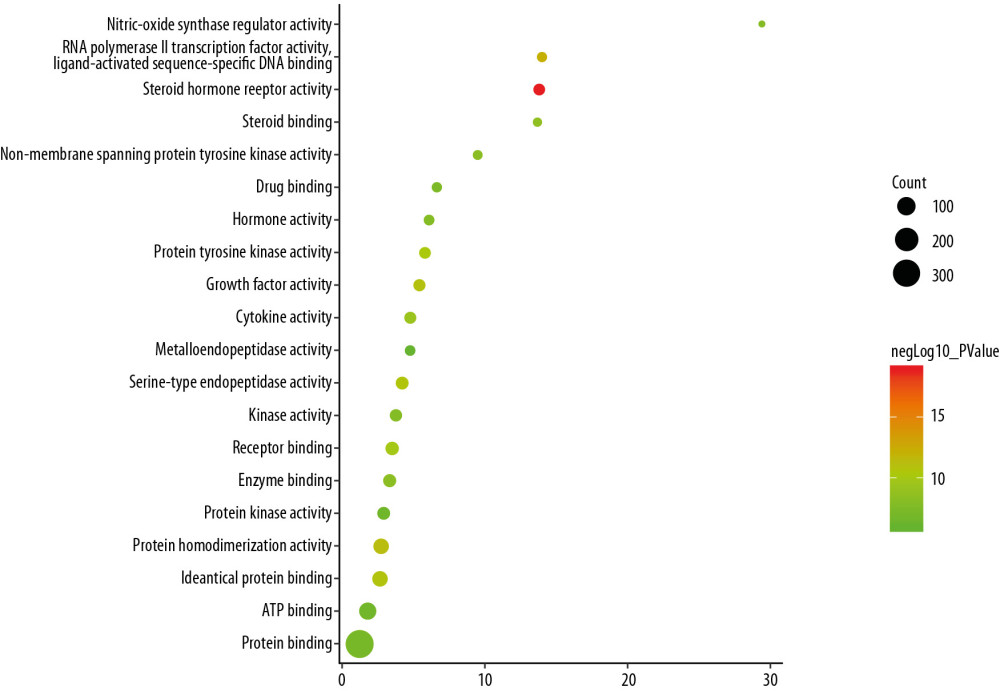 Figure 7. Bubble chart of molecular function (X-axis stand for fold enrichment).
Figure 7. Bubble chart of molecular function (X-axis stand for fold enrichment). 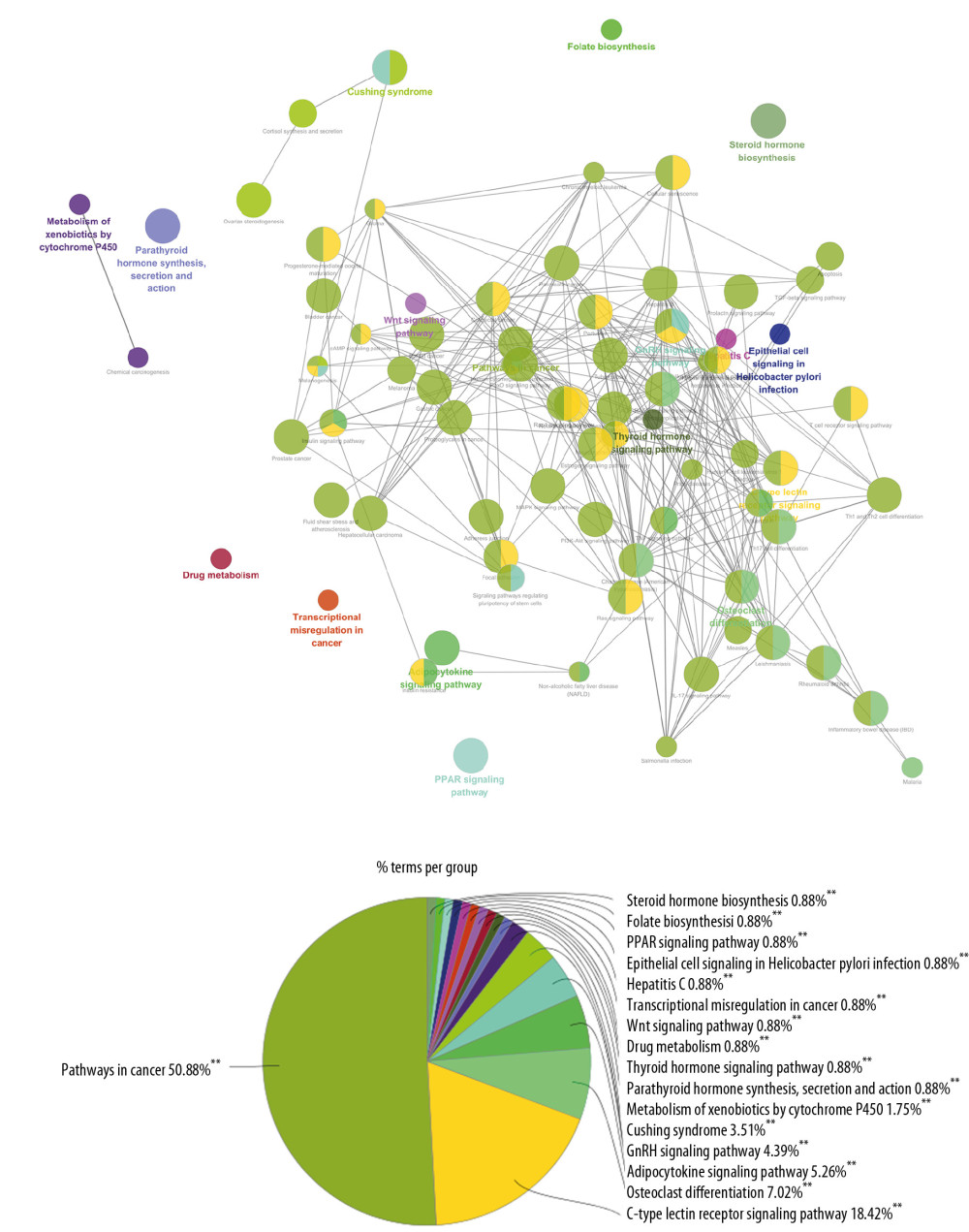 Figure 8. The results of KEGG pathway enrichment analysis.
Figure 8. The results of KEGG pathway enrichment analysis. 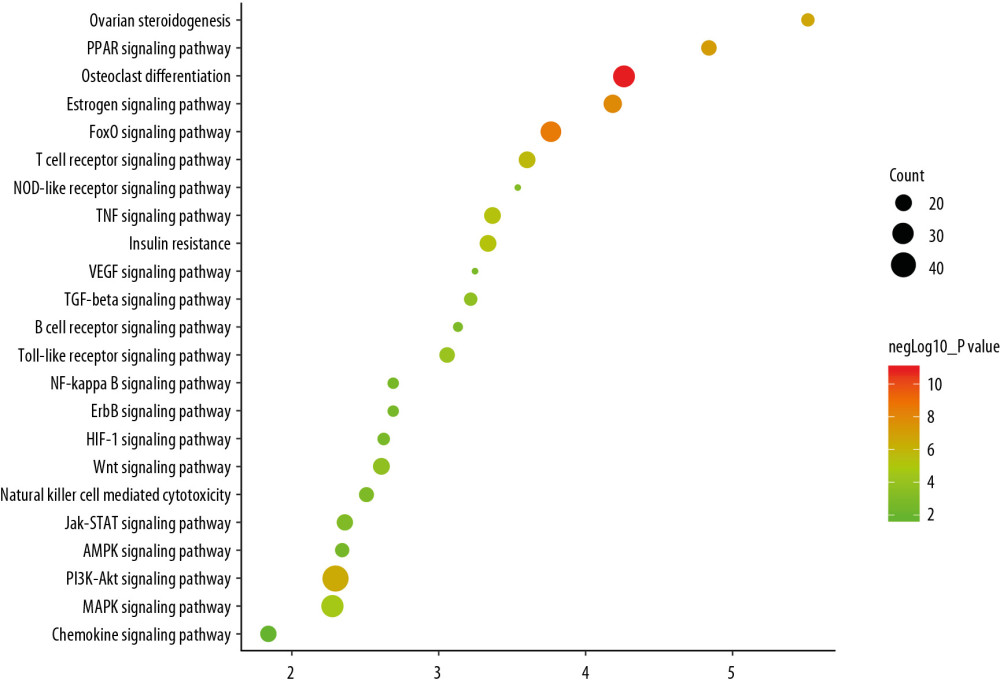 Figure 9. Bubble chart of signaling pathway (X-axis stand for fold enrichment).
Figure 9. Bubble chart of signaling pathway (X-axis stand for fold enrichment). ![Signaling pathways [A: Wnt signaling pathways adapted from KEGG (hsa04310); B: NF-κB signaling pathways adapted from KEGG (hsa04064). The icariin targets were marked in red; the OP genes were marked in yellow; the Icariin-OP target was marked in orange].](https://jours.isi-science.com/imageXml.php?i=medscimonit-26-e924699-g010.jpg&idArt=924699&w=1000) Figure 10. Signaling pathways [A: Wnt signaling pathways adapted from KEGG (hsa04310); B: NF-κB signaling pathways adapted from KEGG (hsa04064). The icariin targets were marked in red; the OP genes were marked in yellow; the Icariin-OP target was marked in orange].
Figure 10. Signaling pathways [A: Wnt signaling pathways adapted from KEGG (hsa04310); B: NF-κB signaling pathways adapted from KEGG (hsa04064). The icariin targets were marked in red; the OP genes were marked in yellow; the Icariin-OP target was marked in orange]. 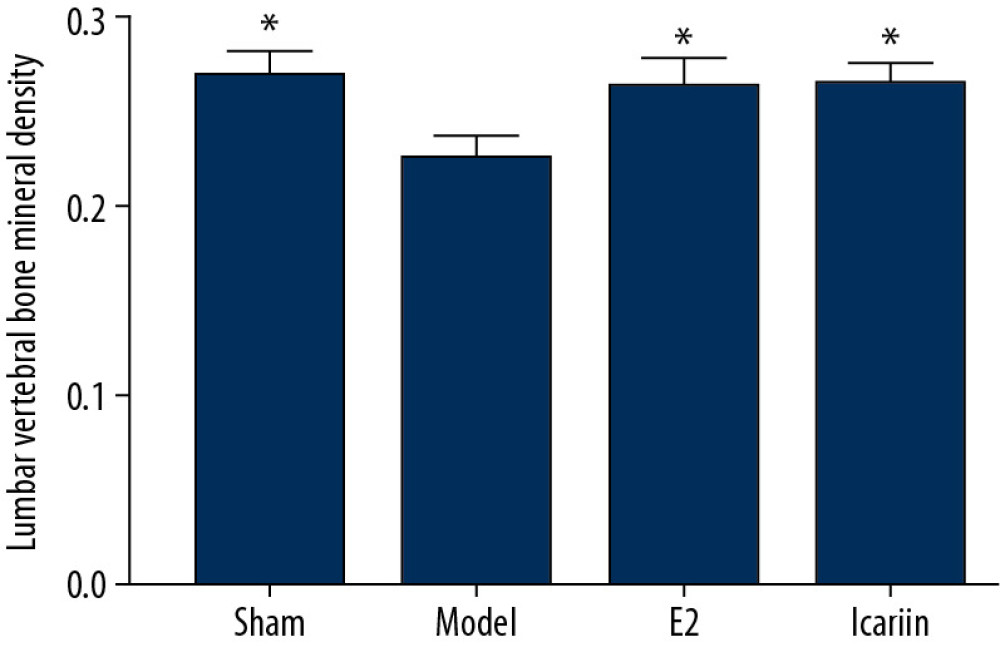 Figure 11. Effect of Icariin on BMD (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).
Figure 11. Effect of Icariin on BMD (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). 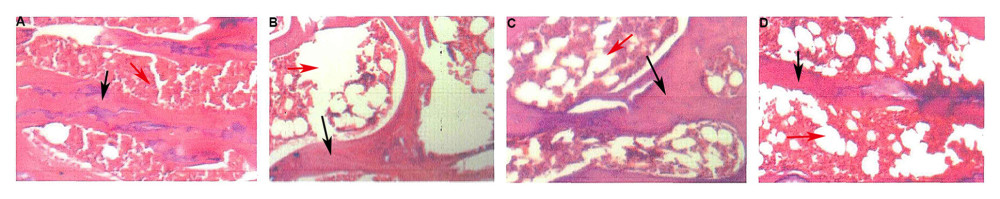 Figure 12. Effect of icariin on morphology of left femur bone in rats (×400, HE staining. After 12 weeks of drug intervention. A: Sham operation group; B: model group; C: E2 group; D: Icariin group. Black arrow indicates trabecular bone; the red arrow indicates the marrow cavity.)
Figure 12. Effect of icariin on morphology of left femur bone in rats (×400, HE staining. After 12 weeks of drug intervention. A: Sham operation group; B: model group; C: E2 group; D: Icariin group. Black arrow indicates trabecular bone; the red arrow indicates the marrow cavity.) 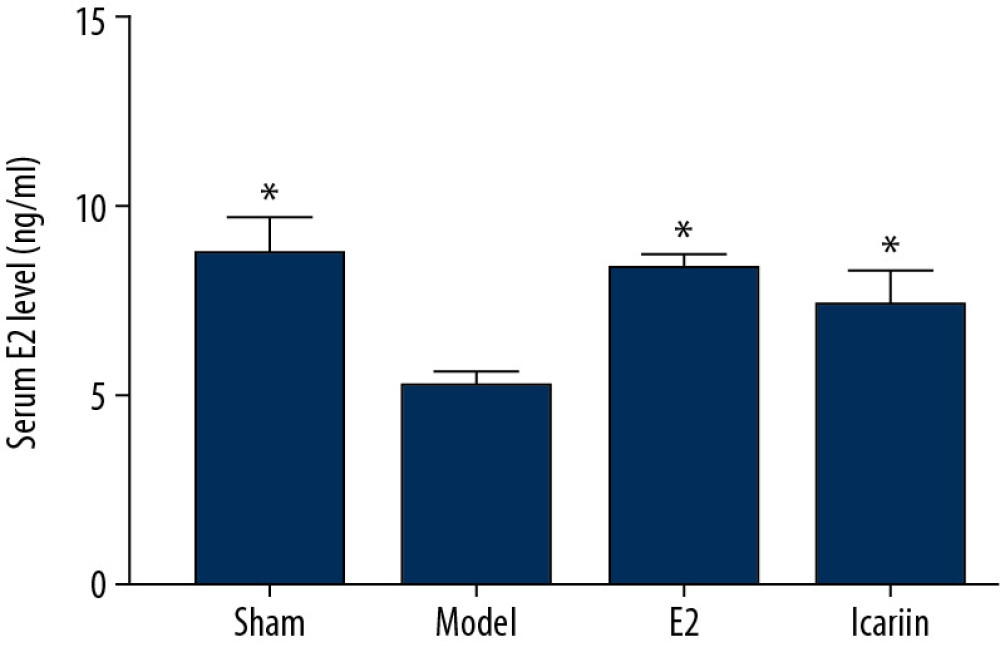 Figure 13. Effect of icariin on serum E2 level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).
Figure 13. Effect of icariin on serum E2 level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). 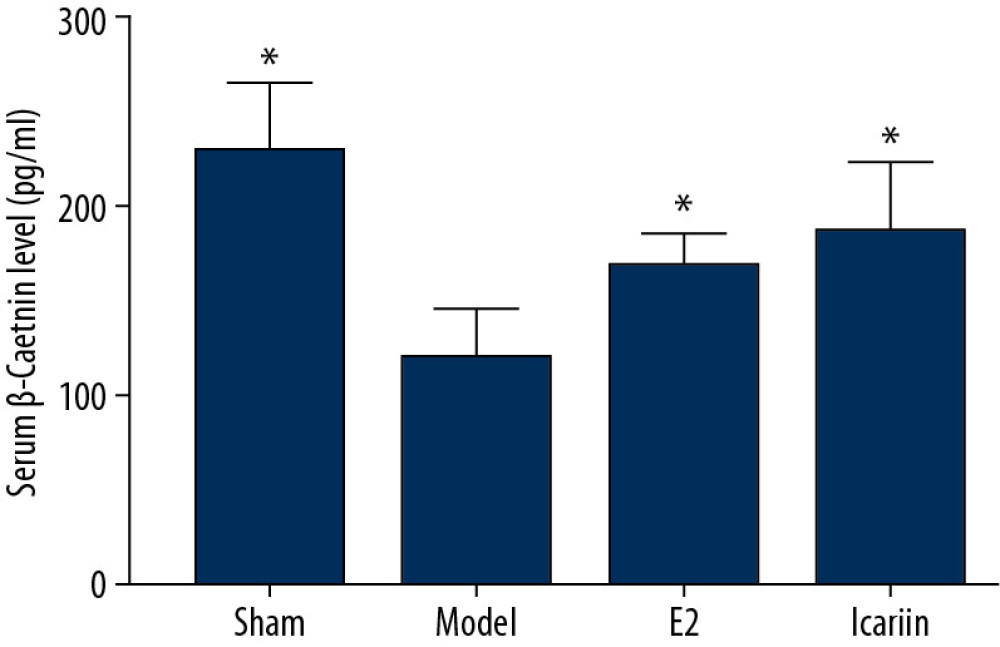 Figure 14. Effect of icariin on serum β-catenin level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).
Figure 14. Effect of icariin on serum β-catenin level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). 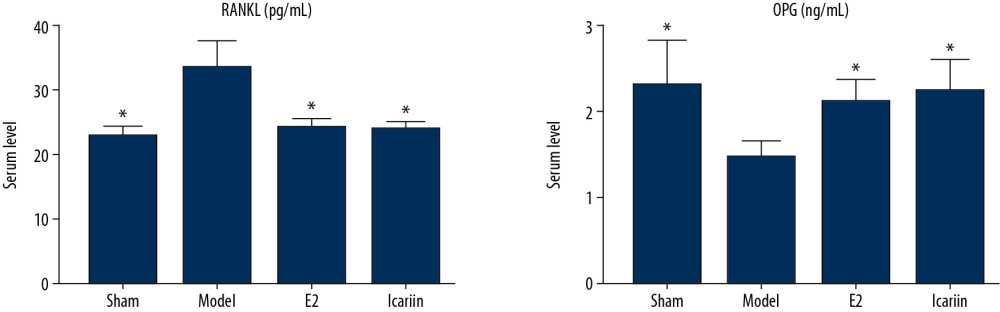 Figure 15. Effect of Icariin on RANKL and OPG level (A: the level of RANK; B: the level of OPG. After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).
Figure 15. Effect of Icariin on RANKL and OPG level (A: the level of RANK; B: the level of OPG. After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). 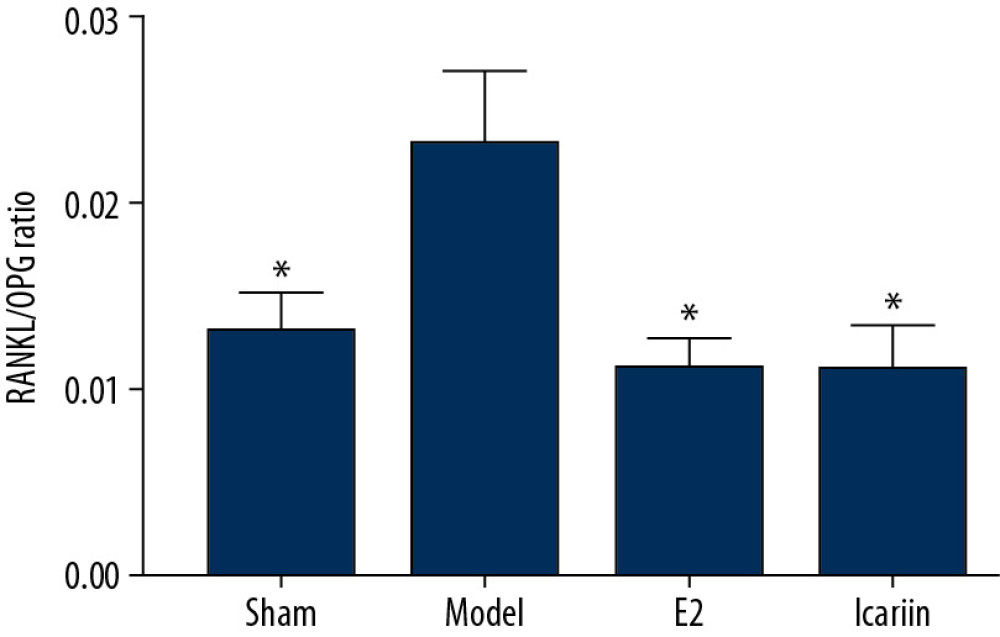 Figure 16. Effect of Icariin on RANKL/OPG ratio (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).
Figure 16. Effect of Icariin on RANKL/OPG ratio (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). References
1. Miller PD, Management of severe osteoporosis: Expert Opin Pharmacother, 2016; 17(4); 473-88
2. Kerschan-Schindl K, Prevention and rehabilitation of osteoporosis: Wien Med Wochenschr, 2016; 166(1–2); 22-27
3. Eastell R, Szulc P, Use of bone turnover markers in postmenopausal osteoporosis: Lancet Diabetes Endocrinol, 2017; 5(11); 908-23
4. Zhou Q, Zhu L, Zhang D, Oxidative stress-related biomarkers in postmenopausal osteoporosis: A systematic review and meta-analyses: Dis Markers, 2016; 2016 7067984
5. Armas LA, Recker RR, Pathophysiology of osteoporosis: New mechanistic insights: Endocrinol Metab Clin North Am, 2012; 41(3); 475-86
6. Cosman F, de Beur SJ, LeBoff MS, Clinician’s guide to prevention and treatment of osteoporosis: Osteoporos Int, 2014; 25(10); 2359-81
7. Black DM, Rosen CJ, Clinical practice. Postmenopausal osteoporosis: N Engl J Med, 2016; 374(3); 254-62
8. Watts NB, Postmenopausal osteoporosis: A clinical review: J Womens Health (Larchmt), 2018; 27(9); 1093-96
9. Wang T, Liu Q, Tjhioe W, Therapeutic potential and outlook of alternative medicine for osteoporosis: Curr Drug Targets, 2017; 18(9); 1051-68
10. Zhang ND, Han T, Huang BK, Traditional Chinese medicine formulas for the treatment of osteoporosis: Implication for antiosteoporotic drug discovery: J Ethnopharmacol, 2016; 189; 61-80
11. Wang Z, Wang D, Yang D, The effect of icariin on bone metabolism and its potential clinical application: Osteoporos Int, 2018; 29(3); 535-44
12. Xu JH, Yao M, Ye J, Bone mass improved effect of icariin for postmenopausal osteoporosis in ovariectomy-induced rats: A meta-analysis and systematic review: Menopause, 2016; 23(10); 1152-57
13. Stéphanou A, Fanchon E, Innominato PF, Ballesta A, Systems biology, systems medicine, systems pharmacology: The what and the why: Acta Biotheor, 2018; 66(4); 345-65
14. Quinney SK, Gullapelli R, Haas DM, Translational systems pharmacology studies in pregnant women: CPT Pharmacometrics Syst Pharmacol, 2018; 7(2); 69-81
15. Xie L, Draizen EJ, Bourne PE, Harnessing big data for systems pharmacology: Annu Rev Pharmacol Toxicol, 2017; 57; 245-62
16. Wang X, Shen Y, Wang S, PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database: Nucleic Acids Res, 2017; 45(W1); W356-60
17. Hamosh A, Scott AF, Amberger JS, Online mendelian inheritance in man (OMIM), a knowledgebase of human genes and genetic disorders: Nucleic Acids Res, 2005; 33; D514-17
18. Stelzer G, Rosen R, Plaschkes I, The GeneCards suite: From gene data mining to disease genome sequence analysis: Curr Protoc Bioinformatics, 2016; 54; 1.30.1-1.30.33
19. Szklarczyk D, Franceschini A, Wyder S, STRING v10: protein-protein interaction networks, integrated over the tree of life: Nucleic Acids Res, 2015; 43; D447-52
20. Bader GD, Hogue CW, An automated method for finding molecular complexes in large protein interaction networks: BMC Bioinformatics, 2003; 4; 2
21. Huang DW, Sherman BT, Lempicki RA, Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources: Nat Protoc, 2009; 4; 44-57
22. Yakar S, Courtland HW, Clemmons D, IGF-1 and bone: New discoveries from mouse models: J Bone Miner Res, 2010; 25(12); 2543-52
23. Liu Z, Mohan S, Yakar S, Does the GH/IGF-1 axis contribute to skeletal sexual dimorphism? Evidence from mouse studies: Growth Horm IGF Res, 2015; 27; 7-17
24. Ashpole NM, Herron JC, Mitschelen MC, IGF-1 regulates vertebral bone aging through sex-specific and time-dependent mechanisms: J Bone Miner Res, 2016; 31(2); 443-54
25. Yan J, Herzog JW, Tsang K, Gut microbiota induce IGF-1 and promote bone formation and growth: Proc Natl Acad Sci USA, 2016; 113(47); E7554-63
26. Cavalcanti FN, Lucas TF, Lazari MF, Porto CS, Estrogen receptor ESR1 mediates activation of ERK1/2, CREB, and ELK1 in the corpus of the epididymis: J Mol Endocrinol, 2015; 54(3); 339-49
27. Wang CL, Tang XY, Chen WQ, Association of estrogen receptor alpha gene polymorphisms with bone mineral density in Chinese women: A meta-analysis: Osteoporos Int, 2007; 18(3); 295-305
28. You L, Chen L, Pan L, Chen JY, New insights into the gene function of osteoporosis: Front Biosci (Landmark Ed), 2013; 18; 1088-97
29. Xiong Y, Zhang Y, Xin N: J Steroid Biochem Mol Biol, 2017; 174; 153-60
30. Huang J, Chen L, IL-1β inhibits osteogenesis of human bone marrow-derived mesenchymal stem cells by activating FoxD3/microRNA-496 to repress Wnt signaling: Genesis, 2017; 55(7); 23040
31. Yuan R, Ma S, Zhu X, Core level regulatory network of osteoblast as molecular mechanism for osteoporosis and treatment: Oncotarget, 2016; 7(4); 3692-701
32. Buyon JP, Dooley MA, Meyer WR, Recommendations for exogenous estrogen to prevent glucocorticoid-induced osteoporosis in premenopausal women with oligo- or amenorrhea: Comment on the american college of rheumatology recommendations for the prevention and treatment of glucocorticoid-induc: Arthritis Rheum, 1997; 40(8); 1548-49
33. Dören M, Estrogen therapy for prevention and treatment of osteoporosis: Maturitas, 2002; 43(1); 53-56
34. Lin C, Shao Y, Zeng C, Blocking PI3K/AKT signaling inhibits bone sclerosis in subchondral bone and attenuates post-traumatic osteoarthritis: J Cell Physiol, 2018; 233(8); 6135-47
35. Wu CM, Chen PC, Li TM, Si-Wu-tang extract stimulates bone formation through PI3K/Akt/NF-κB signaling pathways in osteoblasts: BMC Complement Altern Med, 2013; 13; 277
36. Anaraki PK, Patecki M, Tkachuk S, Urokinase receptor mediates osteoclastogenesis via M-CSF release from osteoblasts and the c-Fms/PI3K/Akt/NF-κB pathway in osteoclasts: J Bone Miner Res, 2015; 30(2); 379-88
37. Rossini M, Gatti D, Adami S, Involvement of WNT/beta-catenin signaling in the treatment of osteoporosis: Calcif Tissue Int, 2013; 93(2); 121-32
38. Guañabens N, Gifre L, Peris P, The role of Wnt signaling and sclerostin in the pathogenesis of glucocorticoid-induced osteoporosis: Curr Osteoporos Rep, 2014; 12(1); 90-97
39. Walsh MC, Choi Y, Biology of the RANKL-RANK-OPG system in immunity, bone, and beyond: Front Immunol, 2014; 5; 511
40. Pivonka P, Zimak J, Smith DW, Theoretical investigation of the role of the RANK–RANKL–OPG system in bone remodeling: J Theor Biol, 2010; 262(2); 306-16
41. Sun X, Cao Z, Zhang Q, Aluminum trichloride inhibits osteoblast mineralization via TGF-β1/Smad signaling pathway: Chem Biol Interact, 2016; 244; 9-15
42. An J, Yang H, Zhang Q, Natural products for treatment of osteoporosis: The effects and mechanisms on promoting osteoblast-mediated bone formation: Life Sci, 2016; 147; 46-58
43. Lin J, Zhu J, Wang Y, Chinese single herbs and active ingredients for postmenopausalosteoporosis: From preclinical evidence to action mechanism: Biosci Trends, 2017; 11(5); 496-506
44. Kim B, Lee KY, Park B, Icariin abrogates osteoclast formation through the regulation of the RANKL-mediated TRAF6/NF-κB/ERK signaling pathway in Raw264.7 cells: Phytomedicine, 2018; 51; 181-90
45. Xu Q, Chen G, Liu X, Icariin inhibits RANKL-induced osteoclastogenesis via modulation of the NF-κB and MAPK signaling pathways: Biochem Biophys Res Commun, 2019; 508(3); 902-6
46. Du Sell CD, Nelson ER, Wang X, The endogenous selective estrogen receptor modulator 27-hydroxycholesterol is a negative regulator of bone homeostasis: Endocrinology, 2010; 151(8); 3675-85
47. Arnal JF, Fontaine C, Abot A: Steroids, 2013; 78(6); 576-82
48. Colaianni G, Sun L, Zaidi M, Zallone A, Oxytocinandbone: Am J Physiol Regul Integr Comp Physiol, 2014; 307(8); R970-77
49. Greenhill C, Bone: High27-hydroxycholesteroldecreasesBMD: Nat Rev Endocrinol, 2010; 6(9); 474
50. Zhai YK, Guo X, Pan YL, A systematic review of the efficacy and pharmacological profile of Herba Epimedii in osteoporosis therapy: Pharmazie, 2013; 68(9); 713-22
51. Zhang G, Qin L, Shi Y, Epimedium-derived phytoestrogen flavonoids exert beneficial effect on preventing bone loss in late postmenopausal women: A 24-month randomized, double-blind and placebo-controlled trial: J Bone Miner Res, 2007; 22(7); 1072-79
52. Westendorf JJ, Kahler RA, Schroeder TM, Wnt signaling in osteoblasts and bone diseases: Gene, 2004; 341(27); 19-39
53. Rawadi G, Roman-Roman S, Wnt signalling pathway: A new target for the treatment of osteoporosis: Expert Opin Ther Targets, 2005; 9(5); 1063-77
54. Mandelin J, Li TF, Liljeström M, Imbalance o fRANKL/RANK/OPG system in interface tissue in loosening of total hipreplacement: J Bone Joint Surg Br, 2003; 85(8); 1196-201
55. Suzuki H, Mochizuki A, Yoshimura K, Bropirimine inhibitsost eoclast differentiation through production of interferon-β: Biochem Biophys Res Commun, 2015; 467(1); 146-51
56. Kostenuik PJ, Osteoprotegerin and RANKL regulate bone resorption, density, geometry and strength: Curr Opin Pharmacol, 2005; 5(6); 618-25
57. Kenkre JS, Bassett J, The bone remodelling cycle: Ann Clin Biochem, 2018; 55(3); 308-27
58. Qi S, He J, Zheng H, Chen C, Lan S, Icariin prevents diabetes-induced bone loss in rats by reducing blood glucose and suppressing bone turnover: Molecules, 2019; 24(10); 1871 :
59. Zheng H, Qi S, Chen C, Salidroside improves bone histomorphology and prevents bone loss in ovariectomized diabetic rats by upregulating the OPG/RANKL ratio: Molecules, 2018; 23(9); 2398
60. Qi S, He J, Han H: Food Funct, 2019; 10(9); 5350-60
61. Chen G, Wang C, Wang J, Antiosteoporotic effect of icariin in ovariectomized rats is mediated via the Wnt/β-catenin pathway: Exp Ther Med, 2016; 12(1); 279-87
Figures
Figure 1. The research process.Figure 2. Venn diagram of icariin potential targets and OP genes.Figure 3. Icariin-OP PPI Network (Pink circles stand for icariin targets. Blue circles stand for OP genes; Purple circles stand for Icariin-OP targets).Figure 4. The results of GO enrichment analysis.Figure 5. Bubble chart of biological processes (X-axis stand for fold enrichment).Figure 6. Bubble chart of cell components (X-axis stand for fold enrichment).Figure 7. Bubble chart of molecular function (X-axis stand for fold enrichment).Figure 8. The results of KEGG pathway enrichment analysis.Figure 9. Bubble chart of signaling pathway (X-axis stand for fold enrichment).Figure 10. Signaling pathways [A: Wnt signaling pathways adapted from KEGG (hsa04310); B: NF-κB signaling pathways adapted from KEGG (hsa04064). The icariin targets were marked in red; the OP genes were marked in yellow; the Icariin-OP target was marked in orange].Figure 11. Effect of Icariin on BMD (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).Figure 12. Effect of icariin on morphology of left femur bone in rats (×400, HE staining. After 12 weeks of drug intervention. A: Sham operation group; B: model group; C: E2 group; D: Icariin group. Black arrow indicates trabecular bone; the red arrow indicates the marrow cavity.)Figure 13. Effect of icariin on serum E2 level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).Figure 14. Effect of icariin on serum β-catenin level (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).Figure 15. Effect of Icariin on RANKL and OPG level (A: the level of RANK; B: the level of OPG. After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05).Figure 16. Effect of Icariin on RANKL/OPG ratio (After 12 weeks of drug intervention. n=8, * Compared with model group, P<0.05). In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387