16 March 2021: Lab/In Vitro Research
Micro-RNA-451 Reduces Proliferation of B-CPAP Human Papillary Thyroid Cancer Cells by Downregulating Expression of Activating Transcription Factor 2
Mei-Feng Zhang1ABDE*, Zhe-Wei Fei1CE, Lei Huang1DEDOI: 10.12659/MSM.929774
Med Sci Monit 2021; 27:e929774






Abstract
BACKGROUND: MicroRNAs (miRNAs) are novel biomarkers that are important in tumorigenesis and cancer treatment resistance. miR-451 is expressed in human papillary thyroid carcinoma (PTC) tissues and is associated with tumor progression. This study investigated the molecular mechanism associated with the effects of miR-451 on B-CPAP human PTC cells in vitro.
MATERIAL AND METHODS: Binding of miRNAs to the 3’ untranslated region (3’UTR) of messenger RNA (mRNA) was determined with a luciferase reporter assay. miRNAs and plasmids were transfected into human PTC B-CPAP cells with Lipofectamine 2000 Transfection Reagent. Cell viability was tested with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay. The levels of miRNAs and mRNA were determined with quantitative polymerase chain reaction and protein levels were analyzed with immunoblotting.
RESULTS: miR-451 bound to wild-type but not mutant 3’-UTR of activating transcription factor 2 (ATF2). MiR-451 mimics inhibited the growth of B-CPAP cells and reduced mRNA and protein levels in ATF2, whereas miR-451 inhibitors promoted the growth of B-CPAP cells and increased mRNA and protein levels in ATF2.
CONCLUSIONS: miR-451 directly bound to the 3’UTR of ATF2, decreased mRNA and protein levels in ATF2, and inhibited growth of B-CPAP cells. Our findings suggest that miR-451 may be a potential therapeutic target for PTC.
Keywords: Activating Transcription Factor 2, in vitro, carcinogenesis, Carcinoma, Papillary, HEK293 Cells, Thyroid Cancer, Papillary, Thyroid Gland
Background
Thyroid cancer is the most common thyroid malignancy in humans and its incidence is increasing worldwide [1–3]. Thyroid cancers are classified as papillary (PTC), follicular, medullary, or anaplastic. PTC accounts for about 85% of all thyroid cancers [1]. With rapid advances in next-generation sequencing and precision medicine, there is an urgent need to identify novel biomarkers for the diagnosis and treatment of and prognostication in PTC.
MicroRNAs (miRNAs) are novel biomarkers that play crucial roles in cancer initiation, progression, and maintenance [4]. They are endogenous, small, non-coding RNAs [5,6]. miRNAs can bind to the 3′ untranslated region (3′UTR) of the target gene messenger RNA (mRNA) to inhibit protein translation by targeting mRNA degradation or translational repression [7]. miRNAs regulate expression of more than 60% of protein-coding genes and an miRNA can function as either an oncogene or tumor suppressor in a target gene- or tissue-dependent manner [8,9]. In a previous study, microRNA-451 (miR-451) was shown to be expressed in human PTC tissues and was associated with tumor progression [10]. Therefore, the present study aimed to investigate the molecular mechanism associated with the effects of miR-451 in B-CPAP human PTC cells in vitro.
Here, we predicted the potential target genes for miR-451 using a bioinformatics database, validated that prediction with a luciferase reporter assay, and analyzed the role of miR-451 in the survival of the PTC cells.
Material and Methods
REAGENTS:
Control miRNAs, miR-451 mimics, and miR-451 inhibitors were obtained from GenePharma Technology (Suzhou, Jiangsu Province, China). A TaqMan™ MicroRNA Reverse Transcription Kit (no. 4366596), TaqMan™ Universal Master Mix II (no. 4440040), hsa-miR-451 assay (assay ID: 001141), RNU44 assay (assay ID: 001094), High-Capacity cDNA Reverse Transcription kit (no. 4368813), TRIzol reagent (no. 15596018), Lipofectamine 2000 Transfection Reagent (no. 11668019), MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide) (no. M6494), anti-activating transcription factor 2 (ATF2) (no. MA5-15807), and anti-β-actin (no. MA1-140) antibodies were purchased from Thermo Fisher Scientific (Waltham, Massachusetts, United States). HEK-293 cells (no. GNHu43) and a human PTC cell line B-CPAP (no. SCSP-543) were purchased from the Stem Cell Bank at the Chinese Academy of Sciences (Shanghai, China).
CELL CULTURE:
B-CPAP cells were maintained in RPMI 1640 medium (Gibco, Carlsbad, California, United States) containing 10% fetal bovine serum (FBS, Gibco). HEK-293 cells were cultured in minimum essential medium (Gibco) containing 10% FBS. The cells were maintained at 37°C in a 5% CO2 atmosphere.
CELL TRANSFECTION:
The cells were seeded in 6- or 96-well plates and miRNA was transfected into cells with Lipofectamine™ 2000 according to the manufacturer’s instructions. The transfected cells were cultured for an additional 48 h, followed by further treatments.
3-(4,5-DIMETHYLTHIAZOL-2-YL)-2,5-DIPHENYL-2H-TETRAZOLIUM BROMIDE ASSAY:
The cells were plated into 96-well plates at a density of 2×103 cells/well. At 48 h after transfection, the cells were incubated with MTT reagent for 4 h at 37°C, followed by replacement with dimethyl sulfoxide (Sigma-Aldrich, Munich, Germany) to dissolve solid residues. Absorbance at 570 nm was read using a spectrophotometer.
QUANTITATIVE POLYMERASE CHAIN REACTION:
TRIzol reagent was used to extract total cellular RNA. For miRNA, total miRNA was converted to complementary DNA (cDNA) using the TaqMan MicroRNA Reverse Transcription Kit. For mRNA, total RNA was reversed to complementary DNA (cDNA) using the High-Capacity cDNA Reverse Transcription Kit. Quantitative polymerase chain reaction (qPCR) analyses of miRNA and mRNA levels were performed with the ABI 7900HT fast real-time PCR system (Applied Biosystems, Inc., Grand Island, New York, United States). Reactions were performed in triplicate. The qPCR data were analyzed with SDS software version 2.2 (Applied Biosystems) and relative miRNA and mRNA levels were normalized with RNU44 and ACTB, respectively, as internal controls. The primer and miRNA sequences are listed in Table 1.
IMMUNOBLOTTING:
Total protein from cells was prepared using RIPA buffer (Thermo Fisher Scientific), followed by protein concentration determination with the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific). The primary antibodies were mouse anti-ATF2 antibody (1: 200 dilution) and anti-β-actin antibody (1: 1000 dilution). The secondary antibody was goat anti-mouse immunoglobulin G conjugated to horseradish peroxidase (1: 1000 dilution). The protein bands were detected with enhanced chemiluminescence (GE Healthcare, Princeton, New Jersey, United States). β-actin served as the loading control to normalize ATF2.
LUCIFERASE REPORTER ASSAY:
The wild-type (WT) and mutant (MUT) 3′UTR of ATF2 were synthesized with Genscript (Nanjing, China) and inserted downstream of the luciferase reporter vector pGL3 (Promega, Madison, Wisconsin, United States). The HEK-293 cells were co-transfected with miR-451 mimics and luciferase reporter plasmids containing either WT or MUT ATF2 3′UTR. At 48 h after transfection, total protein was extracted for luciferase intensity measurement using the Dual Luciferase Reporter assay system (Promega).
STATISTICAL ANALYSES:
Data were analyzed with GraphPad Prism software version 5.0 (GraphPad Software, San Diego, California, United States) and the SPSS statistical package, version 16.0 for Windows (SPSS, Inc., Chicago, Illinois, United States). Data are presented as means±standard deviations (SDs).
Results
MIR-451 INHIBITED GROWTH OF B-CPAP CELLS:
B-CPAP cells were transfected with control miRNA (control group), miR-451 mimics (mimics group), or miR-451 inhibitor (inhibitors group). Compared to the control group, miR-451 levels in the mimics group were significantly increased (P<0.05) (Figure 1A), and in the inhibitors group were significantly decreased (P<0.05) (Figure 1B). MTT assays showed that miR-451 mimics significantly inhibited the growth of B-CPAP cells (P<0.05), whereas the miR-451 inhibitor significantly promoted cell growth (P<0.05; Figure 1C).
MIR-451 BOUND TO THE 3′UTR OF ATF2:
To explore the role of miR-451 in suppression of B-CPAP cell growth, we predicted the potential gene targets of miR-451 with the TargetScanHuman database (website: http://www.targetscan.org/vert_72/). We found that the ATF2 gene was a candidate target for miR-451. To support this prediction, we used the luciferase reporter assay. We constructed WT and MUT 3′UTR of ATF2 luciferase reporter plasmids (Figure 2A) and transfected them into HEK-293 cells together with miR-451 mimics. In the cells transfected with WT 3′UTR, miR-451 significantly reduced the luciferase activity, whereas in the cells transfected with MUT 3′UTR, miR-451 had no significant effect on luciferase activity (Figure 2B).
MIR-451 SUPPRESSED MRNA LEVELS OF ATF2 IN B-CPAP CELLS:
To confirm that miR-451 targets ATF2, B-CPAP cells were transfected with control miRNAs (control group), miR-451 mimics (mimics group), or miR-451 inhibitor (inhibitors group). After 48 h, ATF2 mRNA levels were assessed with qPCR and miR-451 mimics were found to significantly reduce ATF2 (P<0.05) (Figure 3A). In contrast, miR-451 inhibitor significantly increased ATF2 (P<0.001) (Figure 3B). Moreover, correlation analysis showed that miR-451 levels were inversely correlated with ATF2 mRNA levels (spearman r=−0.8596, P<0.0001; Figure 3C).
MIR-451 REDUCED PROTEIN LEVELS OF ATF2 IN B-CPAP CELLS:
We further assessed ATF2 protein levels by immunoblotting following transfection of B-CPAP cells with control miRNAs, miR-451 mimics, or miR-451 inhibitors. Consistent with the alteration of ATF2 mRNA levels (Figure 3), the protein levels of ATF2 were significantly increased in B-CPAP cells transfected with miR-451 mimics (Figure 4A, 4C), whereas they were significantly decreased in cells transfected with miR-451 inhibitors (Figure 4B, 4D).
Discussion
We previously reported that miR-451 is downregulated in PTC tissues and can serve as a potential biomarker for PTC diagnosis [10]; however, the underlying molecular mechanisms of miR-451 in the maintenance of PTC are largely unknown. In the present study, we demonstrated that miR-451 directly bound to the 3′UTR of
Growing evidence has shown that aberrant miRNA expression is associated with the initiation, development, progression, and maintenance of most cancers [4,6]. Previous studies have demonstrated that miR-451 is downregulated in various types of cancer, including gastric cancer [11–13], bladder cancer [14], esophageal carcinoma [15], hepatocellular carcinoma [16], osteosarcoma [17,18], and glioblastoma [19], indicating that miR-451 may serve as a tumor suppressor. However, little is known about miR-451 expression and its relationship with the progression and treatment resistance of PTC. It has been reported that miR-451 is downregulated in PTC tissues and is associated with lymph node metastasis [10]. To investigate the effect of miR-451 on PTC cells, we treated B-CPAP cells with miR-451 mimics or inhibitors and observed that miR-451 inhibited the growth of B-CPAP cells, suggesting that miR-451 is a tumor suppressor in PTC.
The present study had several limitations. First, only B-CPAP cell lines were used, so more PTC cell lines should be used to validate the findings. Second, in addition to
Conclusions
In summary, our results demonstrated that miR-451 suppresses mRNA and protein levels in
Figures
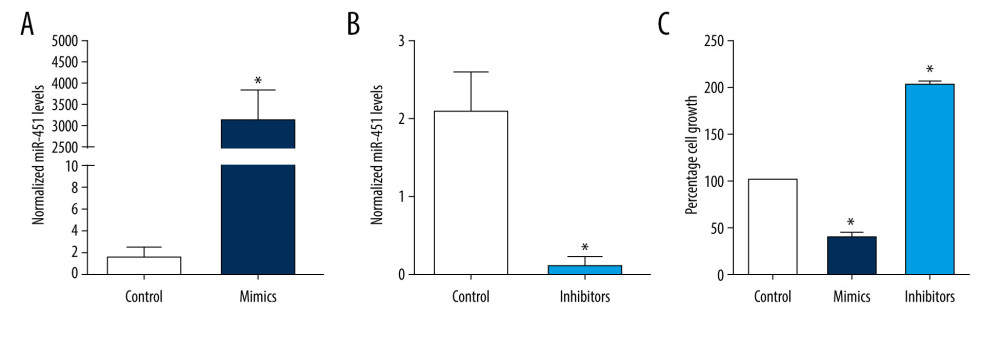 Figure 1. miR-451 inhibited growth of B-CPAP cells. Control microRNA (miRNA) (control, A, B), miR-451 mimics (mimics, A), or miR-451 inhibitors (inhibitors, B) were transfected into B-CPAP cells. After 48 h, miR-451 levels were detected with quantitative polymerase chain reaction. (C) B-CPAP cells were transfected with control miRNAs (control), miR-451 mimics (mimics), or miR-451 inhibitors (inhibitors). Cell viability was measured by with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay and normalized to the control group. * P<0.05 versus the control group.
Figure 1. miR-451 inhibited growth of B-CPAP cells. Control microRNA (miRNA) (control, A, B), miR-451 mimics (mimics, A), or miR-451 inhibitors (inhibitors, B) were transfected into B-CPAP cells. After 48 h, miR-451 levels were detected with quantitative polymerase chain reaction. (C) B-CPAP cells were transfected with control miRNAs (control), miR-451 mimics (mimics), or miR-451 inhibitors (inhibitors). Cell viability was measured by with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay and normalized to the control group. * P<0.05 versus the control group. 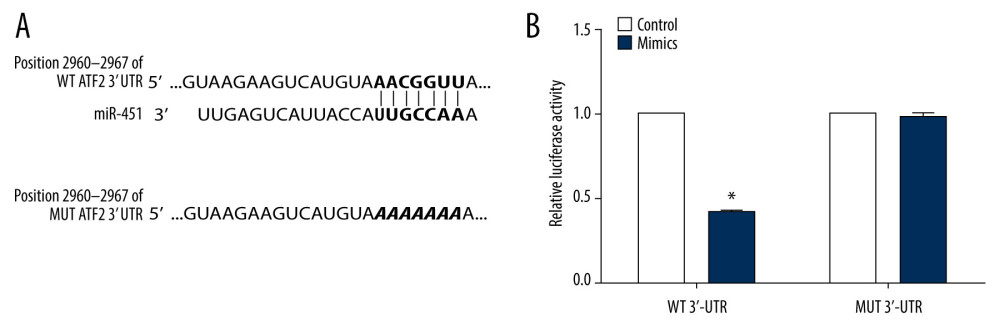 Figure 2. miR-451 directly bound to the 3′UTR of ATF2. (A) The upper panel shows predicted binding sites of miR-451 and ATF2 3′ untranslated region (3′UTR); the bottom panel shows the sequence of MUT ATF2 3′UTR. (B) Using a luciferase reporter assay, HEK-293 cells were co-transfected with miR-451 mimics and reporter plasmid DNA carrying either wild-type (WT) ATF2 3′UTR (WT 3′UTR) or mutant (MUT) ATF2 3′UTR (MUT 3′UTR). Luciferase activity was measured at 48 h after transfection and normalized to the WT 3′-UTR group. *P<0.05 versus the WT 3′UTR group.
Figure 2. miR-451 directly bound to the 3′UTR of ATF2. (A) The upper panel shows predicted binding sites of miR-451 and ATF2 3′ untranslated region (3′UTR); the bottom panel shows the sequence of MUT ATF2 3′UTR. (B) Using a luciferase reporter assay, HEK-293 cells were co-transfected with miR-451 mimics and reporter plasmid DNA carrying either wild-type (WT) ATF2 3′UTR (WT 3′UTR) or mutant (MUT) ATF2 3′UTR (MUT 3′UTR). Luciferase activity was measured at 48 h after transfection and normalized to the WT 3′-UTR group. *P<0.05 versus the WT 3′UTR group. 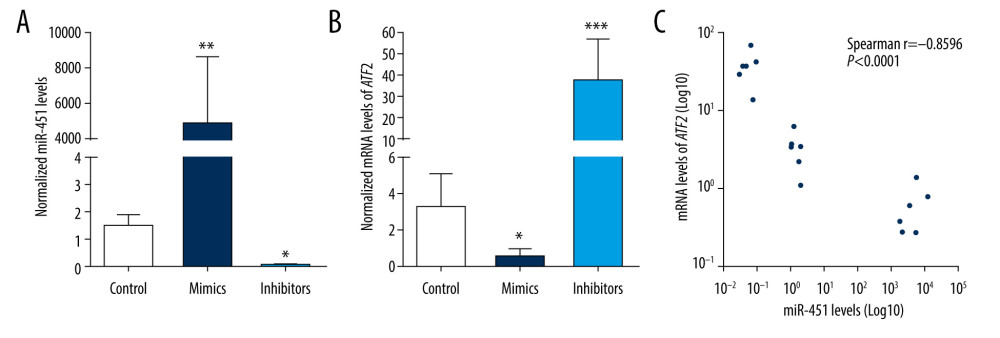 Figure 3. miR-451 negatively regulated mRNA levels of ATF2 in B-CPAP cells. (A, B) B-CPAP cells were transfected with control microRNA (control), miR-451 mimics (mimics) or miR-451 inhibitor (inhibitors) and incubated for 48 h. miR-451 levels (A) and messenger RNA levels of ATF2 (B) were detected with quantitative polymerase chain reaction. (C) Correlation of miR-451 levels and ATF2 mRNA. * P<0.05, ** P<0.01, and *** P<0.001 versus the control group.
Figure 3. miR-451 negatively regulated mRNA levels of ATF2 in B-CPAP cells. (A, B) B-CPAP cells were transfected with control microRNA (control), miR-451 mimics (mimics) or miR-451 inhibitor (inhibitors) and incubated for 48 h. miR-451 levels (A) and messenger RNA levels of ATF2 (B) were detected with quantitative polymerase chain reaction. (C) Correlation of miR-451 levels and ATF2 mRNA. * P<0.05, ** P<0.01, and *** P<0.001 versus the control group. 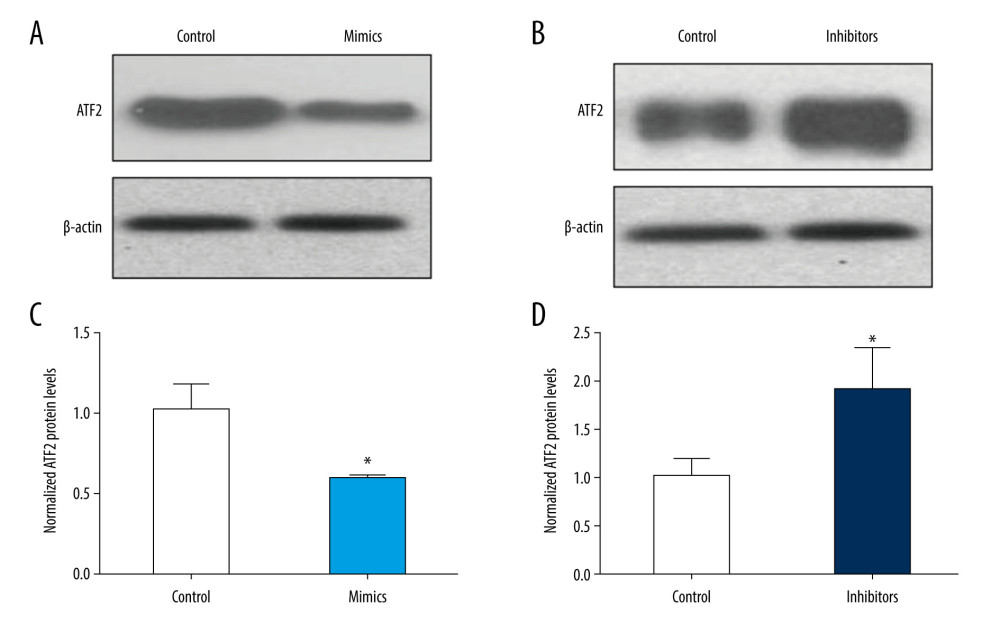 Figure 4. miR-451 negatively regulated protein levels of ATF2 in B-CPAP cells. Control miRNA (control), miR-451 mimics (mimics), or miR-451 inhibitor (inhibitors) were transfected into B-CPAP cells. After 48 h, the protein levels of ATF2 were detected with immunoblotting. (A, B) Representative images of immunoblotting. (C, D) Relative band intensity of the immunoblotting bands (C for A, D for B). Data are shown as the mean±SD; *P<0.05 versus the control group.
Figure 4. miR-451 negatively regulated protein levels of ATF2 in B-CPAP cells. Control miRNA (control), miR-451 mimics (mimics), or miR-451 inhibitor (inhibitors) were transfected into B-CPAP cells. After 48 h, the protein levels of ATF2 were detected with immunoblotting. (A, B) Representative images of immunoblotting. (C, D) Relative band intensity of the immunoblotting bands (C for A, D for B). Data are shown as the mean±SD; *P<0.05 versus the control group. 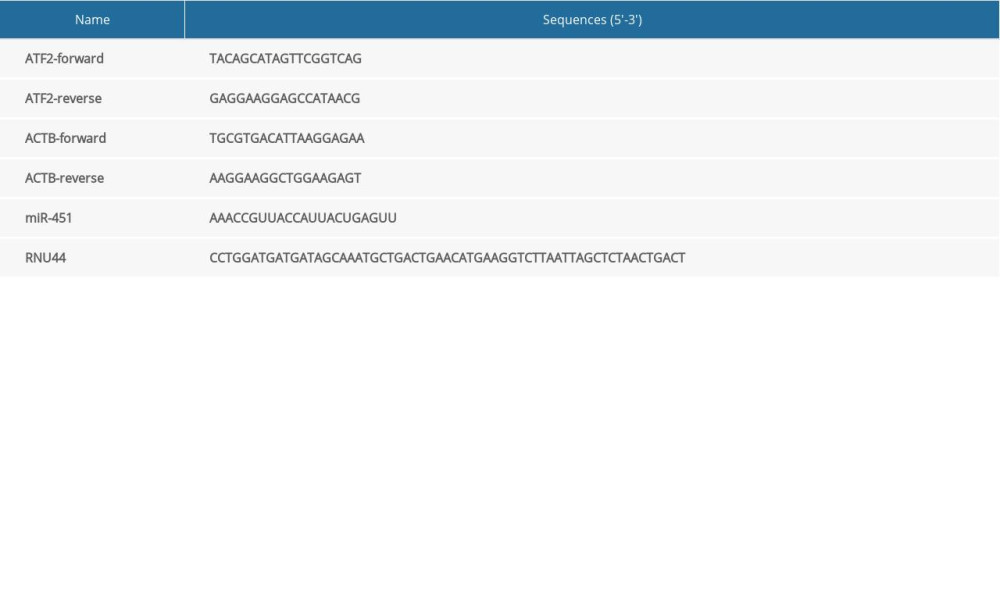
References
1. Chruscik A, Lam AK, Clinical pathological impacts of microRNAs in papillary thyroid carcinoma:A crucial review: Exp Mol Pathol, 2015; 99; 393-98
2. Papp S, Asa SL, When thyroid carcinoma goes bad:A morphological and molecular analysis: Head Neck Pathol, 2015; 9; 16-23
3. Walsh S, Prichard R, Hill AD, Emerging therapies for thyroid carcinoma: Surgeon, 2012; 10; 53-58
4. Lei SL, Zhao H, Yao HL, Regulatory roles of microRNA-708 and microRNA-31 in proliferation, apoptosis and invasion of colorectal cancer cells: Oncol Lett, 2014; 8; 1768-74
5. Li Z, Rana TM, Therapeutic targeting of microRNAs:Current status and future challenges: Nat Rev Drug Discov, 2014; 13; 622-38
6. Hammond SM, An overview of microRNAs: Adv Drug Deliv Rev, 2015; 87; 3-14
7. Sole C, Arnaiz E, MicroRNAs as biomarkers of B-cell lymphoma: Biomark Insights, 2018; 13; 1177271918806840
8. Asghari F, Haghnavaz N, Baradaran B, Tumor suppressor microRNAs:Targeted molecules and signaling pathways in breast cancer: Biomed Pharmacother, 2016; 81; 305-17
9. Svoronos AA, Engelman DM, Slack FJ, OncomiR or tumor suppressor? The duplicity of microRNAs in cancer: Cancer Res, 2016; 76; 3666-70
10. Zhang M, Wu W, Gao M, MicroRNA-451 as a prognostic marker for diagnosis and lymph node metastasis of papillary thyroid carcinoma: Cancer Biomark, 2017; 19; 437-45
11. Shen Y, Gong JM, Zhou LL, MiR-451 as a new tumor marker for gastric cancer: Oncotarget, 2017; 8; 56542-45
12. Su Z, Zhao J, Rong Z, MiR-451, a potential prognostic biomarker and tumor suppressor for gastric cancer: Int J Clin Exp Pathol, 2015; 8; 9154-60
13. Ren C, Chen H, Han C, High expression of miR-16 and miR-451 predicating better prognosis in patients with gastric cancer: J Cancer Res Clin Oncol, 2016; 142; 2489-96
14. Zeng T, Peng L, Chao C, miR-451 inhibits invasion and proliferation of bladder cancer by regulating EMT: Int J Clin Exp Pathol, 2014; 7; 7653-62
15. Zang WQ, Yang X, Wang T, MiR-451 inhibits proliferation of esophageal carcinoma cell line EC9706 by targeting CDKN2D and MAP3K1: World J Gastroenterol, 2015; 21; 5867-76
16. Huang JY, Zhang K, Chen DQ, MicroRNA-451:Epithelial-mesenchymal transition inhibitor and prognostic biomarker of hepatocelluar carcinoma: Oncotarget, 2015; 6; 18613-30
17. Liu SY, Deng SY, He YB, miR-451 inhibits cell growth, migration and angiogenesis in human osteosarcoma via down-regulating IL 6R: Biochem Biophys Res Commun, 2017; 482; 987-93
18. Liu W, Liu SY, He YB, MiR-451 suppresses proliferation, migration and promotes apoptosis of the human osteosarcoma by targeting macrophage migration inhibitory factor: Biomed Pharmacother, 2017; 87; 621-67
19. Alural B, Ayyildiz ZO, Tufekci KU, Erythropoietin promotes glioblastoma via miR-451 suppression: Vitam Horm, 2017; 105; 249-71
20. Wu DS, Chen C, Wu ZJ, ATF2 predicts poor prognosis and promotes malignant phenotypes in renal cell carcinoma: J Exp Clin Cancer Res, 2016; 35; 108
21. Zhang X, Zhang Y, Fan C, Noxin promotes proliferation of breast cancer cells via P38-ATF2 signaling pathway: Tumour Biol, 2017; 39; 1010428317705515
22. Li M, Wu X, Liu N, Silencing of ATF2 inhibits growth of pancreatic cancer cells and enhances sensitivity to chemotherapy: Cell Biol Int, 2017; 41; 599-610
23. Lo Iacono M, Monica V, Vavala T, ATF2 contributes to cisplatin resistance in non-small cell lung cancer and celastrol induces cisplatin resensitization through inhibition of JNK/ATF2 pathway: Int J Cancer, 2015; 136; 2598-609
24. Sun X, Lou L, Zhong K, MicroRNA-451 regulates chemoresistance in renal cell carcinoma by targeting ATF-2 gene: Exp Biol Med (Maywood), 2017; 242; 1299-305
Figures
Figure 1. miR-451 inhibited growth of B-CPAP cells. Control microRNA (miRNA) (control, A, B), miR-451 mimics (mimics, A), or miR-451 inhibitors (inhibitors, B) were transfected into B-CPAP cells. After 48 h, miR-451 levels were detected with quantitative polymerase chain reaction. (C) B-CPAP cells were transfected with control miRNAs (control), miR-451 mimics (mimics), or miR-451 inhibitors (inhibitors). Cell viability was measured by with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay and normalized to the control group. * P<0.05 versus the control group.Figure 2. miR-451 directly bound to the 3′UTR of ATF2. (A) The upper panel shows predicted binding sites of miR-451 and ATF2 3′ untranslated region (3′UTR); the bottom panel shows the sequence of MUT ATF2 3′UTR. (B) Using a luciferase reporter assay, HEK-293 cells were co-transfected with miR-451 mimics and reporter plasmid DNA carrying either wild-type (WT) ATF2 3′UTR (WT 3′UTR) or mutant (MUT) ATF2 3′UTR (MUT 3′UTR). Luciferase activity was measured at 48 h after transfection and normalized to the WT 3′-UTR group. *P<0.05 versus the WT 3′UTR group.Figure 3. miR-451 negatively regulated mRNA levels of ATF2 in B-CPAP cells. (A, B) B-CPAP cells were transfected with control microRNA (control), miR-451 mimics (mimics) or miR-451 inhibitor (inhibitors) and incubated for 48 h. miR-451 levels (A) and messenger RNA levels of ATF2 (B) were detected with quantitative polymerase chain reaction. (C) Correlation of miR-451 levels and ATF2 mRNA. * P<0.05, ** P<0.01, and *** P<0.001 versus the control group.Figure 4. miR-451 negatively regulated protein levels of ATF2 in B-CPAP cells. Control miRNA (control), miR-451 mimics (mimics), or miR-451 inhibitor (inhibitors) were transfected into B-CPAP cells. After 48 h, the protein levels of ATF2 were detected with immunoblotting. (A, B) Representative images of immunoblotting. (C, D) Relative band intensity of the immunoblotting bands (C for A, D for B). Data are shown as the mean±SD; *P<0.05 versus the control group. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387