24 August 2020: Animal Study
Qi-Xian Decoction Upregulated E-cadherin Expression in Human Lung Epithelial Cells and Ovalbumin-Challenged Mice by Inhibiting Reactive Oxygen Species-Mediated Extracellular-Signal-Regulated Kinase (ERK) Activation
Lingling Tang1BCDG*, Linyun Zhu1BCD, Wei Zhang2CDF, Xiaoyan Yang1DE, Qingge Chen1DG, Ziyu Meng1D, Jinjin Liu1E, Yipeng Sun1F, Junsheng Hu1F, Zhenhua Ni3AEG, Xiongbiao Wang1AEGDOI: 10.12659/MSM.922003
Med Sci Monit 2020; 26:e922003






Abstract
BACKGROUND: Loss of the epithelial barrier is characterized by a reduction in E-cadherin expression and is a hallmark of asthma. Qi-xian decoction (QXT) is a Chinese medicinal formula that has been used to effectively treat asthma. This study aimed to investigate the effect of QXT on E-cadherin expression in human lung epithelial 16HBE cells and ovalbumin-challenged mice and to explore the underlying molecular mechanism.
MATERIAL AND METHODS: Ovalbumin (OVA)-induced mice were used as a model of asthma. Real-time PCR and Western blotting were utilized to examine mRNA and protein levels. Lung tissue reactive oxygen species (ROS) levels were evaluated using dichloro-dihydro-fluorescein diacetate (DCFH-DA). Serum superoxide dismutase (SOD) and the total antioxidant capacity (TAOC) were measured via enzyme-linked immunosorbent assay (ELISA)-based analyses. 16HBE cells were utilized to explore the effect of QXT or hydrogen peroxide (H₂O₂) on the expression of E-cadherin in vitro.
RESULTS: We found that QXT treatment increased E-cadherin expression and decreased extracellular-signal-regulated kinase (ERK) phosphorylation levels in the lung tissues of OVA-challenged mice. QXT also downregulated ROS levels and increased serum SOD and TAOC levels in OVA-challenged mice. In vitro studies demonstrated that increased ROS generation induced by H₂O₂ resulted in decreased E-cadherin expression levels in 16HBE cells, which was attenuated by inhibition of ERK signaling. Moreover, the H₂O₂-induced downregulation of E-cadherin expression, increased ROS generation, and ERK activation in 16HBE cells were restored by treatment with QXT water or ethanol extract.
CONCLUSIONS: These data demonstrate that one mechanism by which QXT protects against asthma is to restore E-cadherin expression in vivo and in vitro by inhibiting ROS-mediated ERK activation.
Keywords: Asthma, Cadherins, Extracellular Signal-Regulated MAP Kinases, Lung, Ovalbumin, Phosphorylation
Background
Asthma is a common lung disease and is characterized by chronic airway inflammation; moreover, there is strong evidence that ROS and reactive nitrogen species (RNS) play important roles in asthma [1,2]. Sources of oxidative stress arise from metabolic reactions that use oxygen, and elevated amounts of ROS are released from inflammatory cells that encounter various environmental factors, including air pollutants and cigarette smoke. Excessive ROS and RNS, which are indicators of oxidative stress in response to allergen or pollutant stimulation, have been found in asthma patients [3]. Increased ROS levels play important roles in the pathogenesis of asthma, such as airway epithelial barrier dysfunction, airway inflammation, and airway remodeling [4,5].
E-cadherin is a calcium-dependent adhesion molecule that is expressed on epithelial cells and plays important roles in maintaining epithelial cell junctions [6]. It has recently been demonstrated that E-cadherin expression is reduced in bronchial epithelial cells that were biopsied from asthma patients, and the reduced epithelial E-cadherin, along with catenin or ZO-1, in asthma patients contributes to a defective airway epithelial barrier and an increased influx of eosinophils in the epithelium [7,8]. Furthermore, the loss of E-cadherin-mediated cell–cell contacts is essential for epithelial-to-mesenchymal transition (EMT), and E-cadherin is a potential target for ameliorating asthma severity or progression in humans [9]. E-cadherin is regulated by various factors, such as transforming growth factor β1 (TGF-β1), and ROS is one of the triggers that downregulates E-cadherin [10,11]. Therefore, blocking ROS production and recovering E-cadherin expression is a promising approach in asthma therapy. However, the signaling pathways that are involved in ROS-mediated E-cadherin expression have not been fully elucidated.
The use of traditional Chinese medicine (TCM), a type of alternative medicine, is constantly growing in asthma treatment because of effectiveness and less severe adverse effect [12,13]. For example, Xiao-qing-long-tang (XQLT), a traditional Chinese medicine, has been used to treat patients with bronchial asthma in Asian countries for several centuries.
Qi-xian decoction (Chinese patent: ZL201310158130.3) is a Chinese formula composed of
Material and Methods
ANIMALS:
The procedures for each experiment were approved by the Institutional Animal Care and Use Committee, Putuo Hospital, Shanghai University of Traditional Chinese Medicine. Animal studies were conducted in strict conformity to the established institutional guidelines and the NIH guidelines on the use of experimental animals.
Female BALB/c mice were purchased from Shanghai SLAC Laboratory Animal Co. (Shanghai, China). All mice were maintained in well-controlled animal housing conditions.
PREPARATION OF QXT:
Traditional Chinese herbs were purchased from Shanghai Hua Yu Machinery Co., Shanghai, China. One set of QXT components is listed in Table 1. All components of QXT were evaluated by the Quality Control Department of Shanghai Hua Yu Chinese Herbs Co. according to Chinese medicine component processing standards. In brief, 2 sets of QXT components were boiled with 2 liters of cold water and concentrated to create a boiled water extract (272 mL) and stored at 4°C prior to administration to the mice.
OVA-CHALLENGED MOUSE MODEL OF ASTHMA AND QXT TREATMENT:
Previous studies indicated that female mice were more susceptible to the development of allergic airway inflammation, AHR, and airway remodeling [25–27]. Therefore, female BALB/c mice were used to establish the murine asthma model, as previously described [28]. Twenty-four female BALB/c mice, 6- to 8-weeks-old, were divided into the following 3 groups (8 mice/group): the saline group (mice were challenged and treated with saline), the OVA group (mice were challenged with OVA and treated with saline), and the OVA+QXT group (mice were challenged with OVA and treated with QXT). On days 0 and 14, the OVA and OVA+QXT groups were sensitized via intraperitoneal (i.p.) injection of 100 μg of OVA (Grade V; Sigma, St. Louis, MO, USA) complexed with alum. On days 14, 25, 26, and 27, the OVA group OVA+QXT group were intranasally administered 50 μL of OVA. Between days 28 and 34, the OVA+QXT group was intragastrically (i.g.) administered 0.5 mL QXT (37.5 g/kg/d) once daily, and the saline and OVA groups received 0.5 mL normal saline. On day 35, the mice were euthanized by cervical dislocation; then, the lungs were dissected, and the serum was collected for further analysis.
LUNG ROS ASSAY:
ROS levels in the lung tissue were assayed with DCFH-DA (Nanjin Jiancheng Bioengineering Institute, Nanjing, China). Briefly, 0.2 g of fresh lung tissue was cut into 1-mm3 sections and enzymatically digested at 37°C for 25 min. Digested lung tissue was extruded through a metal strainer followed by a nylon mesh screen to harvest single-cell suspensions. 5×105 cells were resuspended in 1 mL pre-warmed PBS containing 1 μL of DCFH-DA for 30 min at room temperature. We analyzed 300 μL of cell suspension using a microplate reader with excitation wavelength of 500 nm and emission wavelength of 525 nm.
SOD AND TAOC ASSAY:
SOD and TAOC levels were assayed according to the instructions provided by the manufacturer (Nanjin Jiancheng Bioengineering Institute, Nanjing, China). The SOD level was measured at 450 nm, and the TAOC level was measured at 520 nm by a microplate reader.
PREPARATION OF WATER AND ETHANOL EXTRACT OF QXT:
In brief, 2 set of mixed herbs with 2 L cold water were boiled and concentrated to 0.45 L to prepare a boiled water extract, of which 0.27 L concentrated decoction was then used for ethanol extraction (QXT-E) and the remaining 0.18 L was used for water extraction (QXT-W). For ethanol extraction, 0.27 L decoction was adjusted to 1.17L by adding 0.9L 90% ethanol. We let the decoction stand for 24 h, then the supernatant was collected and ethanol was evaporated by vacuum distillation. The remaining solution was freeze-dried and the final weigh of lyophilized powder was 49.1 g. The extraction rate by ethanol was 25.73%. The powders were dissolved in dimethyl sulfoxide (DMSO) for use as ethanol extract of QXT (QXT-E) in
CELL CULTURE:
16HBE cell line (Bogoo Biotech Co., Shanghai, China) cells were cultivated at 37°C and 5% CO2 in RPMI-1640 medium supplemented with 10% FBS and penicillin-streptomycin solution (all from Thermo Fisher Scientific, Inc., Waltham, MA, USA). Cells were treated with H2O2 (40 μM, Grade V, Sigma, St. Louis, MO, U.S.A.), QXT-E or QXT-W (80 μg/ml), or solvent control (DMSO plus water) at 37°C for indicated times.
GENE SILENCING:
Small-interfering RNAs (siRNA) against the human ERK1 (Gene ID: 5595), ERK2 (Gene ID: 5594) were synthesized by RiboBio (Guangzhou, China). Cells were transfected with siRNAs by TransIT-TKO (Mirus, Madison, WI). The SiRNA sequences were:
REAL-TIME QUANTITATIVE PCR (QPCR):
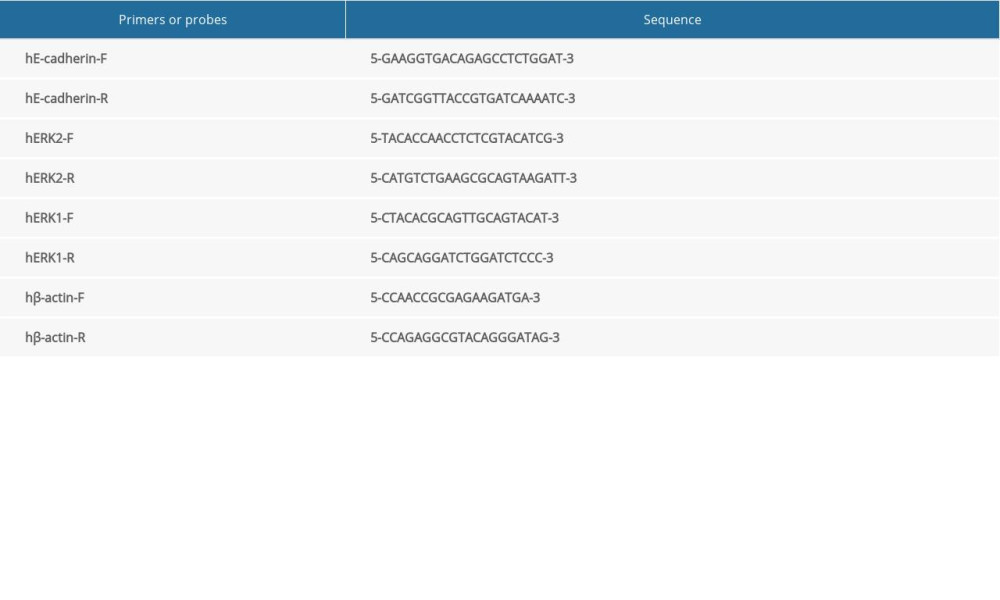
Total RNA was extracted from the lungs or cells using Trizol reagent (TaKaRa, Dalian, China) according to manufacturer’s instructions, and first-strand cDNAs was obtained from the extracted RNA by using the First-Strand Synthesis Kit (Roche, San Francisco, CA, USA). qPCR primers are listed in Table 2. qPCR was carried out using the Universal Master Mix (Roche). The thermocycling conditions were: 94°C for 10 min, 45 cycles of 94°C for 30 s, and 60°C for 1 min. The relative expression levels of target genes were normalized against β-actin and calculated using the comparative 2−ΔΔCt method.
WESTERN BLOT:
Western blot analysis was done according to published methods [28,29]. Briefly, the lung tissues or cells were extracted with cell lysis reagent (Sigma, St. Louis, MO, USA ). Then, protein samples were separated with 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, Bedford, MA, USA). The PVDF membrane was incubated in 5% bovine serum albumin (BSA) for 2 h. After 3 washes with Tris-buffered saline containing Tween-20 (TBST), the PVDF membrane was incubated with E-cadherin, β-actin, P-ERK1/2 and ERK1/2 antibodies (Cell Signaling Technology, Danvers, USA) separately overnight at 4°C. The PVDF membrane was then washed with TBST 3 times followed by incubation with HRP-conjugated secondary antibodies (Cell Signaling Technology, Danvers, USA) for 2 h at room temperature. After 3 washes with TBST, immunoreactive bands were visualized using chemiluminescence detection reagents (Bio-Rad Laboratories, Redmond, USA). The signal intensities of bands were quantified by using ImageJ software.
IMMUNOFLUORESCENCE:
After fixing in 4% paraformaldehyde, the cells were permeabilized with 0.1% Triton X-100 in PBS 3 times (5 min each) and incubated in blocking solution for 30 min at room temperature. After blocking, the cells were washed with PBS 3 times (5 min each) and incubated in anti-human E-cadherin (Cell Signaling Technology, Danvers, USA) overnight at 4°C. Then, the cells were washed 3 times with PBS, incubated for 30 min with Alexa Fluor 488-labeled Goat Anti-Rabbit IgG (Abcam, Cambridge, UK), and washed another 3 times with PBS. Nuclei were stained with 4’, 6-diamidino-2-phenylindole (DAPI) and the sections were visualized using a fluorescence microscope.
STATISTICAL ANALYSIS:
In all the experiments, data are expressed as the mean±standard deviation and were analyzed using SPSS version 21.0 software. Means were compared using the unpaired
Results
QXT INCREASED E-CADHERIN EXPRESSION IN OVA-CHALLENGED MICE:
To investigate the effects of QXT on E-cadherin expression in vivo, BALB/c mice were used to establish a model of asthma. Histopathological examination of the mouse lungs confirmed that airway inflammation and smooth muscle thickening were increased in OVA-challenged mice and attenuated in QXT-treated mice (Figure 1A). Moreover, an increased level of serum IgE (Figure 1B) was observed in the asthmatic mice compared to that of the saline group and was reduced after treatment with QXT. These results confirmed the protective effects of QXT in preventing asthma features in OVA-challenged mice. Furthermore, our results showed that mice that were treated with QXT had significantly increased protein levels of E-cadherin compared to the effects of OVA alone (Figure 1C, 1D), which demonstrated that QXT restored E-cadherin expression in OVA-challenged mice.
QXT INHIBITED ROS GENERATION IN OVA-CHALLENGED MICE:
ROS are one of the triggers of the downregulation of E-cadherin [30]. We thus investigated whether QXT inhibited ROS generation in OVA-challenged mice. Our results showed increased ROS production in the lung tissues of OVA-challenged mice compared to that of the saline group, and treatment with QXT decreased intracellular ROS levels (Figure 2A). Moreover, we found that both serum superoxide dismutase (SOD) and total antioxidant capacity (TAOC) were lower in OVA-challenged mice than in the saline group. However, treatment with QXT increased the levels of serum SOD and TAOC (Figure 2B, C). These results demonstrated that QXT modulated ROS production by increasing SOD activity, which contributes to increased E-cadherin expression in OVA-challenged mice.
:
To further evaluate whether QXT restores E-cadherin expression by regulating ROS, the water or ethanol extraction of QXT was prepared for an in vitro study. 16HBE cells were treated with H2O2 (40 μmol/l) to induce ROS generation. We found that H2O2 stimulation decreased E-cadherin expression in a time-dependent manner in 16HBE cells (Figure 3A–3C). We then investigated the effect of QXT extract on H2O2-mediated E-cadherin expression. As shown in Figure 3D–3H, the H2O2-induced downregulation of E-cadherin at both the mRNA and protein levels was attenuated by QXT extract. Moreover, H2O2 treatment decreased SOD and TAOC levels in 16HBE cells, whereas ROS, SOD, and TAOC levels were increased after treatment with ethanol or water extract of QXT (Figure 3I–3L). Our results demonstrated that H2O2 treatment increased cellular ROS levels and decreased E-cadherin expression, whereas QXT extract alleviated H2O2-mediated E-cadherin downregulation and ROS production in 16HBE cells.
:
We investigated the mechanism by which ROS regulates E-cadherin expression. We observed that the phosphorylation levels of ERK were upregulated by H2O2 treatment (Figure 4A, 4B). To further investigate the roles of ERK activation on H2O2-mediated E-cadherin expression, we inhibited ERK activation by using RNA interference. As shown in Figure 4C–4E, the phosphorylation levels of ERK were reduced by ERK siRNA, and the expression of E-cadherin mRNA and protein was restored in ERK siRNA-treated cells compared with those of control cells after H2O2 treatment (Figure 4F–4H). Furthermore, using an ERK inhibitor, the downregulation of E-cadherin was also attenuated (Figure 4I–4L). Our results indicated that H2O2 treatment inhibited E-cadherin expression in 16HBE cells in an ERK-dependent manner.
ERK ACTIVATION IN OVA-CHALLENGED MICE AND 16HBE CELLS WAS INHIBITED BY QXT OR QXT EXTRACT:
We further examined the phosphorylation levels of ERK1/2 in the lung tissue of OVA-challenged mice. Our results showed that ERK1/2 phosphorylation levels were increased in the OVA-challenged mice compared to those in the control mice. Moreover, treatment with QXT decreased ERK1/2 phosphorylation in vivo (Figure 5A, 5B). Furthermore, the QXT extracts significantly inhibited H2O2-induced ERK phosphorylation in 16HBE cells (Figure 5C, 5D). These results showed that QXT or its extracts inhibit ERK activation in vivo and H2O2-induced ERK activation in vitro.
Discussion
The airway epithelium forms the first line of defense against inhaled environmental insults, which play a critical role in asthma [9]. E-cadherin is an adhesion molecule that is important for maintaining epithelial integrity. Loss of epithelial E-cadherin expression contributes to a defective airway epithelial barrier in asthma, leading to inflammation. Consequently, it may be possible to slow the disease progression of asthma through enhancement of E-cadherin expression. The regulation of E-cadherin expression has been studied in association with EMT in asthma. E-cadherin expression is downregulated by TGF-β and members of the Snail family, such as Sna-1 and Sna-2 [31]. In addition to TGF-β, Wang et al. reported that brahma-related gene-1 (Brg1) inhibited E-cadherin expression in lung epithelial cells by inactivating a critical region within the E-cadherin promoter [32]. In the present study, we demonstrated that increased ROS induced by H2O2 inhibited E-cadherin expression in 16HBE cells. Moreover, downregulation of E-cadherin by H2O2 was involved in ERK pathway activation. Furthermore, the downregulation of E-cadherin and activation of ERK in 16HBE cells and in OVA-challenged mice were counterbalanced by QXT treatment, which was associated with the antioxidative activity of QXT. Our results suggest one potential mechanism whereby oxidative stress signals are transduced to regulate E-cadherin expression, and QXT is a potential effective alternative therapy for restoring E-cadherin expression.
Oxidative stress is a result of an imbalance between ROS generation and ROS removal by various antioxidant molecules [32]. We examined the effect of QXT in regulating antioxidant molecules in OVA-challenged mice. Our results showed increased ROS production in OVA-challenged mice compared to those in the saline group. Treatment with QXT decreased intracellular ROS levels, which indicated the antioxidative activity of QXT
Recently, it has become evident that ROS and cellular oxidative stress are important transducers of intracellular signals that are involved in the development and/or exacerbation of asthma [37,38]. For example, the ERK signaling pathway is activated by ROS, thus inducing airway inflammation and other phenotypes [37,39]. We found that the QXT mice displayed decreased phosphorylation levels of ERK1/2 in lung tissue compared to those of the OVA mice. Moreover, the E-cadherin expression level was also restored by QXT treatment. These findings suggest that QXT inhibits ROS and subsequently regulates ERK1/2 activation and E-cadherin expression. Consistent with our findings, some studies have reported that hydrogen peroxide (H2O2) exposure activates the ERK1/2 signaling pathway in lung adenocarcinoma cells or cardiac fibroblasts [40,41].
By using 2 different extraction methods, we found that both ethanol and water extracts of QXT showed antioxidative effects
Conclusions
Our findings demonstrated that QXT treatment increased E-cadherin expression in OVA-challenged mice by inhibiting ROS production and ERK activation. Consistently, an
Figures
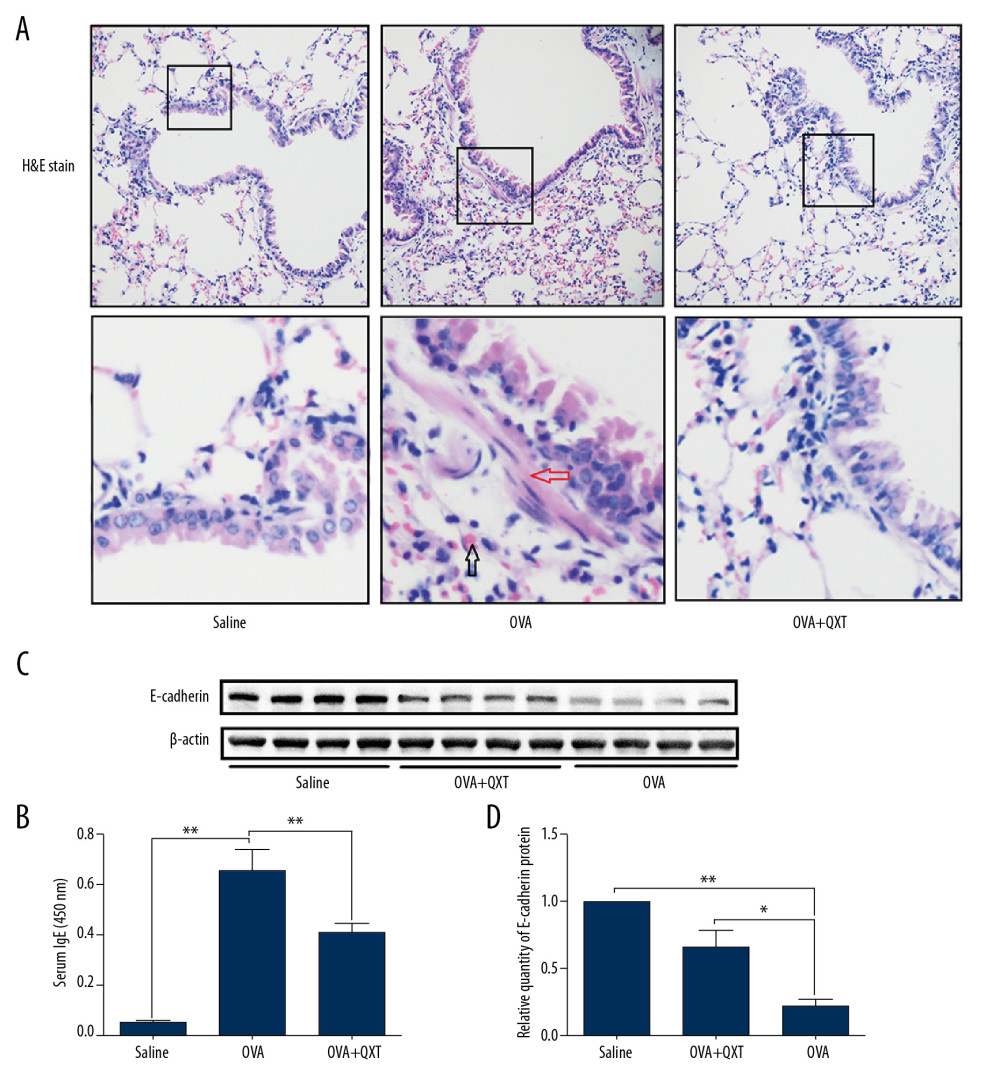 Figure 1. QXT increased E-cadherin expression in OVA-challenged mice. (A) The lung tissues of OVA-challenged mice were examined with hematoxylin and eosin (×100). Infiltration of eosinophils was shown by black arrows and smooth muscle thickening was shown by red arrows (n=8). (B) Serum IgE levels were estimated using ELISA (n=8). (C) E-cadherin levels were determined by Western blot analysis (n=8). (D) E-cadherin protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.
Figure 1. QXT increased E-cadherin expression in OVA-challenged mice. (A) The lung tissues of OVA-challenged mice were examined with hematoxylin and eosin (×100). Infiltration of eosinophils was shown by black arrows and smooth muscle thickening was shown by red arrows (n=8). (B) Serum IgE levels were estimated using ELISA (n=8). (C) E-cadherin levels were determined by Western blot analysis (n=8). (D) E-cadherin protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01. 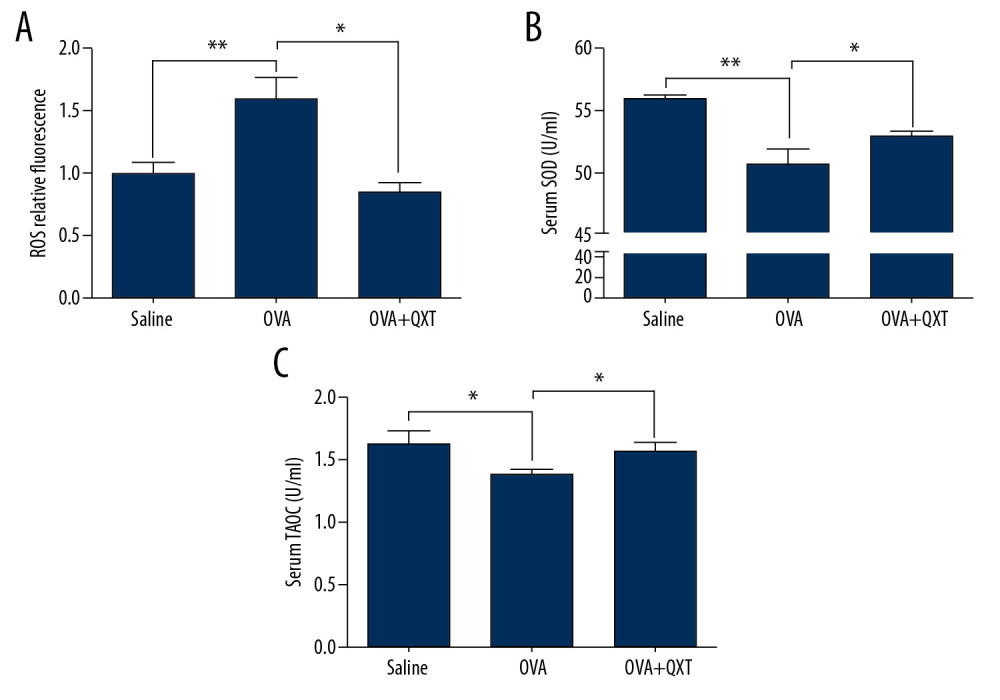 Figure 2. QXT inhibited ROS generation in OVA-challenged mice. (A) ROS levels in the lung tissue were determined using DCFH-DA and measured on a microplate reader (n=8). (B, C) SOD and TAOC serum levels in each group were determined. The SOD level was measured at 450 nm and the TAOC level was measured at 520 nm (n=8). Data are shown as the mean±standard, * P<0.05, ** P<0.01.
Figure 2. QXT inhibited ROS generation in OVA-challenged mice. (A) ROS levels in the lung tissue were determined using DCFH-DA and measured on a microplate reader (n=8). (B, C) SOD and TAOC serum levels in each group were determined. The SOD level was measured at 450 nm and the TAOC level was measured at 520 nm (n=8). Data are shown as the mean±standard, * P<0.05, ** P<0.01. 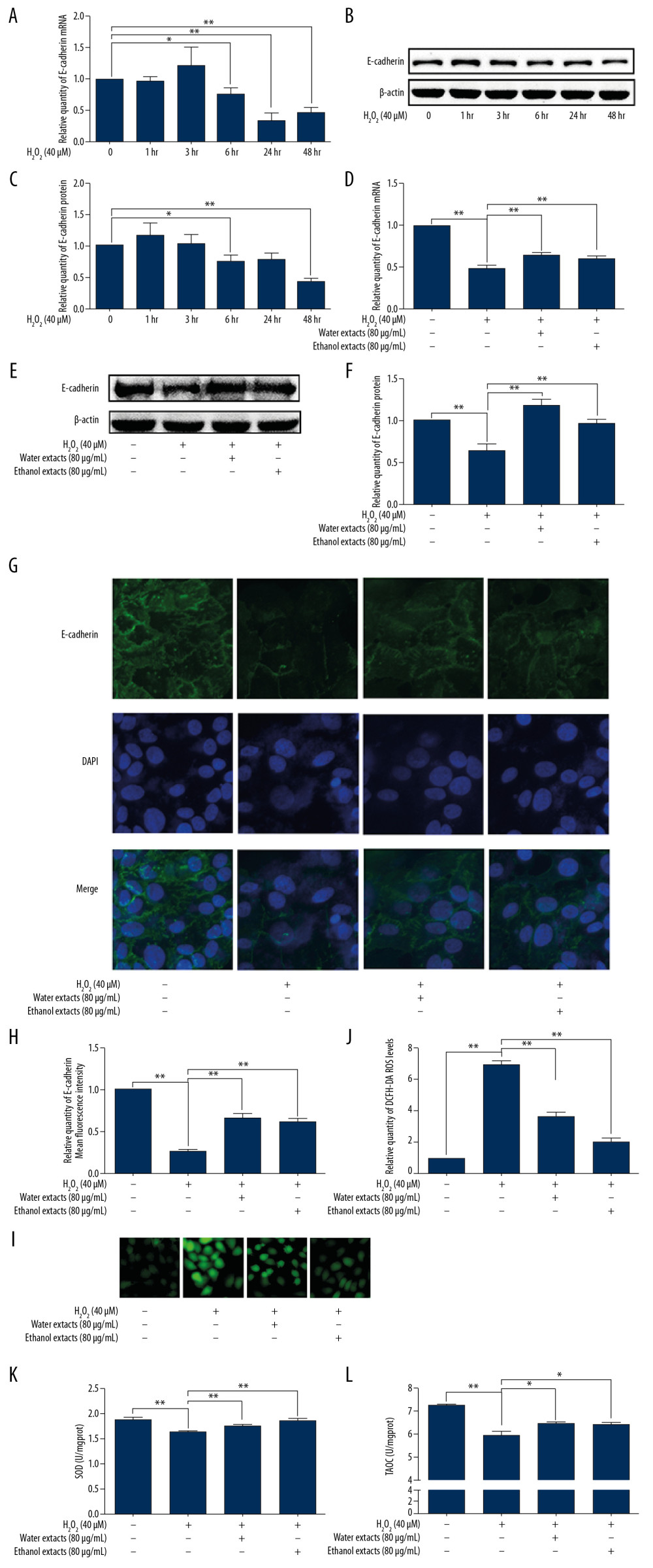 Figure 3. QXT extraction attenuated ROS-mediated E-cadherin downregulation in human lung epithelial cells. (A–C) Cultured 16HBE cells were incubated with H2O2 (40 μM) for different times. The expression level of E-cadherin was determined via qPCR (n=3, A) and Western blotting (n=3, B, C). (D–L) Cultured 16HBE cells were incubated with H2O2 or treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h or 72 h. The expression level of E-cadherin was determined via qPCR (n=3, D), Western blotting (n=3, E, F), and immunofluorescence (n=3, G, H). ROS level was determined via DCFH-DA and observed with a fluorescence microscope (n=3, I, J). The levels of SOD and TAOC were detected by colorimetry (n=3, K, L). The protein levels of E-cadherin were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.
Figure 3. QXT extraction attenuated ROS-mediated E-cadherin downregulation in human lung epithelial cells. (A–C) Cultured 16HBE cells were incubated with H2O2 (40 μM) for different times. The expression level of E-cadherin was determined via qPCR (n=3, A) and Western blotting (n=3, B, C). (D–L) Cultured 16HBE cells were incubated with H2O2 or treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h or 72 h. The expression level of E-cadherin was determined via qPCR (n=3, D), Western blotting (n=3, E, F), and immunofluorescence (n=3, G, H). ROS level was determined via DCFH-DA and observed with a fluorescence microscope (n=3, I, J). The levels of SOD and TAOC were detected by colorimetry (n=3, K, L). The protein levels of E-cadherin were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01. 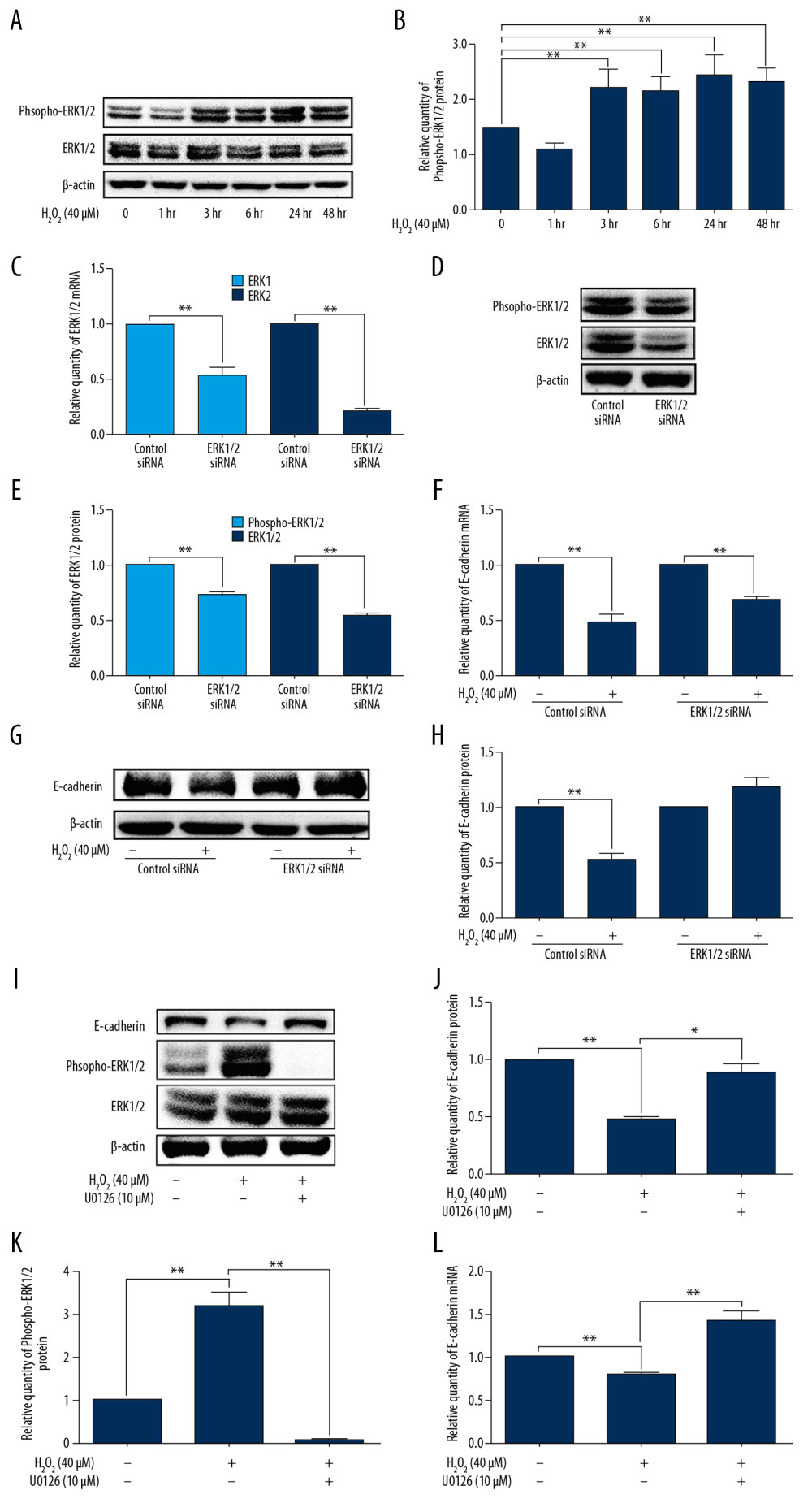 Figure 4. ROS reduced E-cadherin expression in ERK-dependent manner in 16HBE cells. (A, B) 16HBE cells were incubated with H2O2 (40 μM) for indicated times (0, 1, 3, 6, 24, and 48h), the protein levels of phospho-ERK and ERK were measured by using Western blotting (n=3). (C–H) 16HBE cells were transfected by ERK siRNA for 24h and then treated with H2O2 (40μM) for 48 h. The expression level of ERK1/2 (n=3, C–E) and E-cadherin (n=3, F–H) was determined via qPCR and Western blotting. (I–L) 16HBE cells were treated with U0126 (10 μM) plus H2O2 (40 μM) for 48 h. The expression level of ERK1/2 (n=3, I, K) and E-cadherin (n=3, I, J, L) was determined via qPCR and Western blotting. The protein levels of E-cadherin, phospho-ERK, and ERK were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.
Figure 4. ROS reduced E-cadherin expression in ERK-dependent manner in 16HBE cells. (A, B) 16HBE cells were incubated with H2O2 (40 μM) for indicated times (0, 1, 3, 6, 24, and 48h), the protein levels of phospho-ERK and ERK were measured by using Western blotting (n=3). (C–H) 16HBE cells were transfected by ERK siRNA for 24h and then treated with H2O2 (40μM) for 48 h. The expression level of ERK1/2 (n=3, C–E) and E-cadherin (n=3, F–H) was determined via qPCR and Western blotting. (I–L) 16HBE cells were treated with U0126 (10 μM) plus H2O2 (40 μM) for 48 h. The expression level of ERK1/2 (n=3, I, K) and E-cadherin (n=3, I, J, L) was determined via qPCR and Western blotting. The protein levels of E-cadherin, phospho-ERK, and ERK were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01. 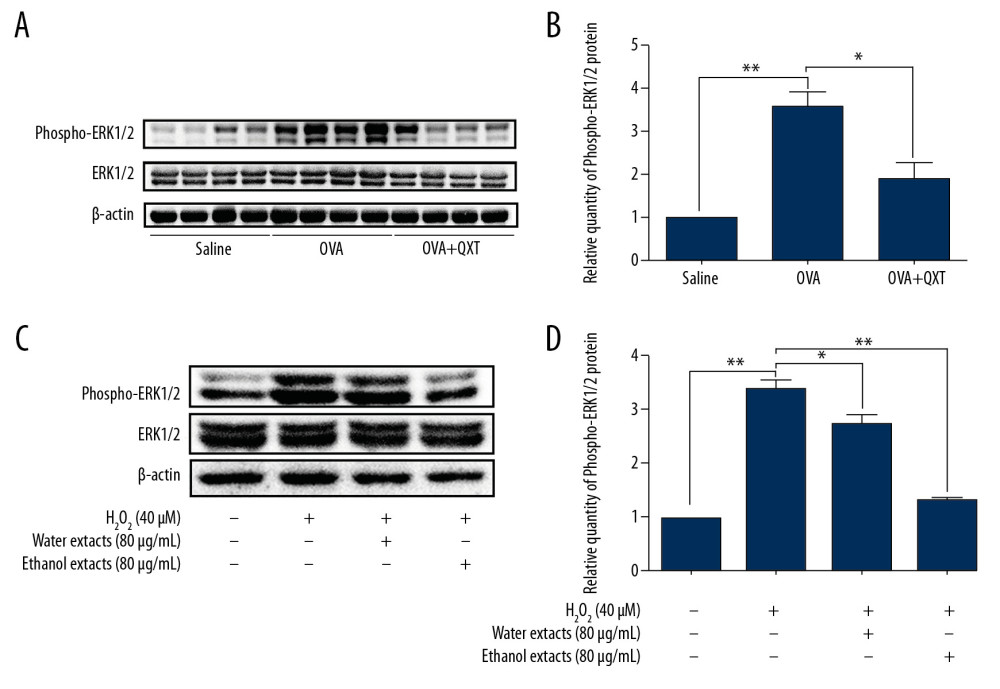 Figure 5. QXT attenuated ERK activation in OVA-challenged mice and human lung epithelial cells. (A, B) ERK1/2 phosphorylation levels were examined by Western blot (n=8). (C, D) 16HBE cells were treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h. The protein levels of phospho-ERK and ERK were detected by Western blotting (n=3). (B, D) Phospho-ERK protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.
Figure 5. QXT attenuated ERK activation in OVA-challenged mice and human lung epithelial cells. (A, B) ERK1/2 phosphorylation levels were examined by Western blot (n=8). (C, D) 16HBE cells were treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h. The protein levels of phospho-ERK and ERK were detected by Western blotting (n=3). (B, D) Phospho-ERK protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01. 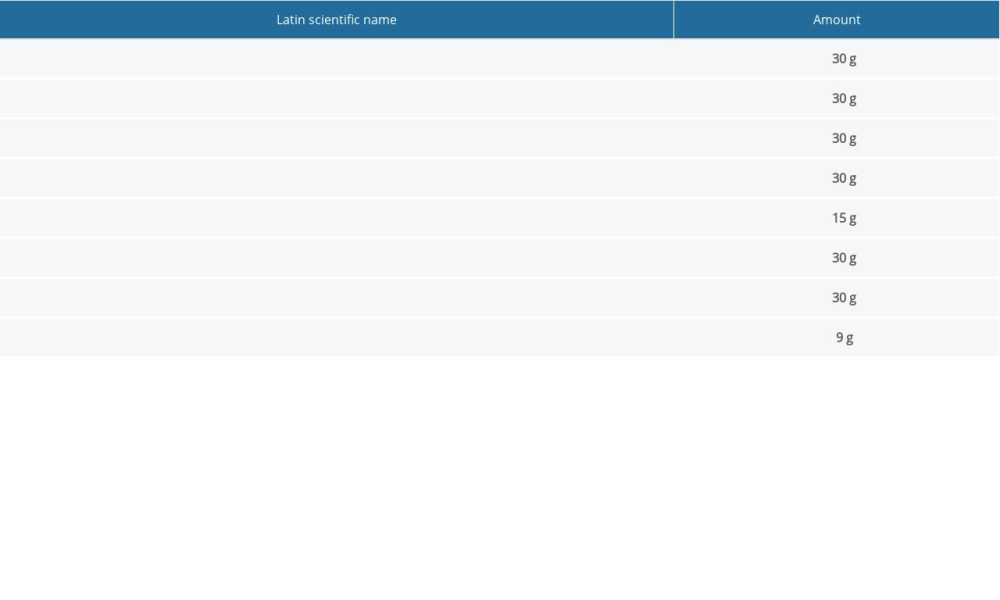
References
1. Sahiner UM, Birben E, Erzurum S, Oxidative stress in asthma: World Allergy Organ J, 2011; 4; 151-58
2. Phaniendra A, Jestadi DB, Periyasamy L, Properties, sources, targets, and their implication in various diseases: Indian J Clin Biochem, 2015; 30; 11-26
3. Ekmekci OB, Donma O, Ekmekci H, Plasma paraoxonase activities, lipoprotein oxidation, and trace element interaction in asthmatic patients: Biol Trace Elem Res, 2006; 111; 41-52
4. Birben E, Sahiner UM, Sackesen C, Oxidative stress and antioxidant defense: World Allergy Organ J, 2012; 5; 9-19
5. Unger BL, Ganesan S, Comstock AT, Nod-like receptor X-1 is required for rhinovirus-induced barrier dysfunction in airway epithelial cells: J Virol, 2014; 88; 3705-18
6. Heijink IH, Kies PM, Kauffman HF, Down-regulation of E-cadherin in human bronchial epithelial cells leads to epidermal growth factor receptor-dependent Th2 cell-promoting activity: J Immunol, 2007; 178; 7678-85
7. de Boer WI, Sharma HS, Baelemans SM, Altered expression of epithelial junctional proteins in atopic asthma: Possible role in inflammation: Can J Physiol Pharmacol, 2008; 86; 105-12
8. Hackett TL, Singhera GK, Shaheen F, Intrinsic phenotypic differences of asthmatic epithelium and its inflammatory responses to respiratory syncytial virus and air pollution: Am J Respir Cell Mol Biol, 2011; 45; 1090-100
9. Heijink IH, Nawijn MC, Hackett TL, Airway epithelial barrier function regulates the pathogenesis of allergic asthma: Clin Exp Allergy, 2014; 44; 620-30
10. Gao H, Wu X, Simon L, Antioxidants maintain E-cadherin levels to limit Drosophila prohemocyte differentiation: PLoS One, 2014; 9; e107768
11. Cui J, Zhao H, Wang C: Oxid Med Cell Longev, 2015; 2015 358396
12. Asher BF, Seidman MD, Reddy WD, Integrative medical approaches to allergic rhinitis: Curr Opin Otolaryngol Head Neck Surg, 2015; 23; 221-25
13. Chen X, Luo Y, Wang M, Wuhu decoction regulates dendritic cell autophagy in the treatment of respiratory syncytial virus (RSV)-induced mouse asthma by AMPK/ULK1 signaling pathway: Med Sci Monit, 2019; 25; 5389-400
14. Kao ST, Lin CS, Hsieh CC: Allergy, 2001; 56; 1164-71
15. Kao ST, Yeh TJ, Hsieh CC, The effects of Ma-Xing-Gan-Shi-Tang on respiratory resistance and airway leukocyte infiltration in asthmatic guinea pigs: Immunopharmacol Immunotoxicol, 2001; 23; 445-58
16. Chan CK, Kuo ML, Shen JJ, Ding Chuan Tang, a Chinese herb decoction, could improve airway hyper-responsiveness in stabilized asthmatic children: A randomized, double-blind clinical trial: Pediatr Allergy Immunol, 2006; 17; 316-22
17. Ma CH, Ma ZQ, Fu Q, Ma Huang Tang ameliorates asthma though modulation of Th1/Th2 cytokines and inhibition of Th17 cells in ovalbumin-sensitized mice: Chin J Nat Med, 2014; 12; 361-66
18. Lai Y, Ni Z, Li ZEffect of Qixianqingming granule on airway inflammation in a mouse model of asthma: Shanghai Medical Journal, 2014; 37; 1037-40 [in Chinese]
19. Sun L, Chen Q, Ni ZMechanism of Qixiantang decoction in the threatment of asthma mice: Herald of Med, 2014; 36; 23-28 [in Chinese]
20. Sze SC, Tong Y, Ng TB, Herba Epimedii: Anti-oxidative properties and its medical implications: Molecules, 2010; 15; 7861-70
21. Shahzad M, Shabbir A, Wojcikowski K: Curr Drug Targets, 2016; 17; 1331-40
22. Hu Y, Sun B, Liu K, Icariin attenuates high-cholesterol diet induced atherosclerosis in rats by inhibition of inflammatory response and p38 MAPK signaling pathway: Inflammation, 2016; 39; 228-36
23. Qian W, Cai X, Qian Q, Astragaloside IV modulates TGF-beta1-dependent epithelial-mesenchymal transition in bleomycin-induced pulmonary fibrosis: J Cell Mol Med, 2018; 22; 4354-65
24. Zeng H, Wang Y, Gu Y, Polydatin attenuates reactive oxygen species-induced airway remodeling by promoting Nrf2-mediated antioxidant signaling in asthma mouse model: Life Sci, 2019; 218; 25-30
25. Takeda M, Tanabe M, Ito W, Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma: Respirology, 2013; 18; 797-806
26. Okuyama K, Wada K, Chihara J, Sex-related splenocyte function in a murine model of allergic asthma: Clin Exp Allergy, 2008; 38; 1212-19
27. Zhan X, Zhang W, Sun T, Bulleyaconitine A effectively relieves allergic lung inflammation in a murine asthmatic model: Med Sci Monit, 2019; 25; 1656-62
28. Tang L, Chen Q, Meng Z, Suppression of Sirtuin-1 increases IL-6 expression by activation of the Akt pathway during allergic asthma: Cell Physiol Biochem, 2017; 43; 1950-60
29. Zhu H, Ji W, Dihydroartemisinin ameliorated ovalbumin-induced asthma in mice via regulation of MiR-183C: Med Sci Monit, 2019; 25; 3804-14
30. Wang R, Ding G, Liang W, Role of LOX-1 and ROS in oxidized low-density lipoprotein induced epithelial-mesenchymal transition of NRK52E: Lipids Health Dis, 2010; 9; 120
31. Nawijn MC, Hackett TL, Postma DS, E-cadherin: Gatekeeper of airway mucosa and allergic sensitization: Trends Immunol, 2011; 32; 248-55
32. Wang T, Zou W, Niu C, Brg1 inhibits E-cadherin expression in lung epithelial cells and disrupts epithelial integrity: J Mol Med (Berl), 2017; 95; 1117-26
33. Afonso V, Champy R, Mitrovic D, Reactive oxygen species and superoxide dismutases: role in joint diseases: Joint Bone Spine, 2007; 74(4); 324-29
34. Ahmad A, Shameem M, Husain Q, Relation of oxidant-antioxidant imbalance with disease progression in patients with asthma: Ann Thorac Med, 2012; 7; 226-32
35. El-Sherbeeny NA, Hassan ZA, Ateyya H, Tiron ameliorates oxidative stress and inflammation in a murine model of airway remodeling: Int Immunopharmacol, 2016; 39; 172-80
36. Joseph A, Li Y, Koo HC, Superoxide dismutase attenuates hyperoxia-induced interleukin-8 induction via AP-1: Free Radic Biol Med, 2008; 45; 1143-49
37. Lee YA, Shin MH, Mitochondrial respiration is required for activation of ERK1/2 and caspase-3 in human eosinophils stimulated with hydrogen peroxide: J Investig Allergol Clin Immunol, 2009; 19; 188-94
38. Aravamudan B, Kiel A, Freeman M, Cigarette smoke-induced mitochondrial fragmentation and dysfunction in human airway smooth muscle: Am J Physiol Lung Cell Mol Physiol, 2014; 306; L840-54
39. Kumari A, Singh DK, Dash D, Intranasal curcumin protects against LPS-induced airway remodeling by modulating toll-like receptor-4 (TLR-4) and matrixmetalloproteinase-9 (MMP-9) expression via affecting MAP kinases in mouse model: Inflammopharmacology, 2019; 27; 731-48
40. Gallelli L, Falcone D, Scaramuzzino M, Effects of simvastatin on cell viability and proinflammatory pathways in lung adenocarcinoma cells exposed to hydrogen peroxide: BMC Pharmacol Toxicol, 2014; 15; 67
41. Sano M, Fukuda K, Sato T, ERK and p38 MAPK, but not NF-kappaB, are critically involved in reactive oxygen species-mediated induction of IL-6 by angiotensin II in cardiac fibroblasts: Circ Res, 2001; 89; 661-69
42. Yanhua W, Fuhua W, Zhaohan G, Optimization of extraction process for polysaccharide in salvia miltiorrhiza bunge using response surface methodology: Open Biomed Eng J, 2014; 8; 153-59
43. Liu Y, Huang G, Extraction and derivatisation of active polysaccharides: J Enzyme Inhib Med Chem, 2019; 34; 1690-96
44. Li L, Hou X, Xu R, Research review on the pharmacological effects of astragaloside IV: Fundam Clin Pharmacol, 2017; 31; 17-36
45. Chen Y, Zhang Y, Xu M, Catalpol alleviates ovalbumin-induced asthma in mice: Reduced eosinophil infiltration in the lung: Int Immunopharmacol, 2017; 43; 140-46
46. Bi J, Jiang B, Liu JH: Neurosci Lett, 2008; 442; 224-27
Figures
Figure 1. QXT increased E-cadherin expression in OVA-challenged mice. (A) The lung tissues of OVA-challenged mice were examined with hematoxylin and eosin (×100). Infiltration of eosinophils was shown by black arrows and smooth muscle thickening was shown by red arrows (n=8). (B) Serum IgE levels were estimated using ELISA (n=8). (C) E-cadherin levels were determined by Western blot analysis (n=8). (D) E-cadherin protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.Figure 2. QXT inhibited ROS generation in OVA-challenged mice. (A) ROS levels in the lung tissue were determined using DCFH-DA and measured on a microplate reader (n=8). (B, C) SOD and TAOC serum levels in each group were determined. The SOD level was measured at 450 nm and the TAOC level was measured at 520 nm (n=8). Data are shown as the mean±standard, * P<0.05, ** P<0.01.Figure 3. QXT extraction attenuated ROS-mediated E-cadherin downregulation in human lung epithelial cells. (A–C) Cultured 16HBE cells were incubated with H2O2 (40 μM) for different times. The expression level of E-cadherin was determined via qPCR (n=3, A) and Western blotting (n=3, B, C). (D–L) Cultured 16HBE cells were incubated with H2O2 or treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h or 72 h. The expression level of E-cadherin was determined via qPCR (n=3, D), Western blotting (n=3, E, F), and immunofluorescence (n=3, G, H). ROS level was determined via DCFH-DA and observed with a fluorescence microscope (n=3, I, J). The levels of SOD and TAOC were detected by colorimetry (n=3, K, L). The protein levels of E-cadherin were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.Figure 4. ROS reduced E-cadherin expression in ERK-dependent manner in 16HBE cells. (A, B) 16HBE cells were incubated with H2O2 (40 μM) for indicated times (0, 1, 3, 6, 24, and 48h), the protein levels of phospho-ERK and ERK were measured by using Western blotting (n=3). (C–H) 16HBE cells were transfected by ERK siRNA for 24h and then treated with H2O2 (40μM) for 48 h. The expression level of ERK1/2 (n=3, C–E) and E-cadherin (n=3, F–H) was determined via qPCR and Western blotting. (I–L) 16HBE cells were treated with U0126 (10 μM) plus H2O2 (40 μM) for 48 h. The expression level of ERK1/2 (n=3, I, K) and E-cadherin (n=3, I, J, L) was determined via qPCR and Western blotting. The protein levels of E-cadherin, phospho-ERK, and ERK were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01.Figure 5. QXT attenuated ERK activation in OVA-challenged mice and human lung epithelial cells. (A, B) ERK1/2 phosphorylation levels were examined by Western blot (n=8). (C, D) 16HBE cells were treated with H2O2 (40 μM) plus the water or ethanol extract from QXT (80 μg/mL) for 48 h. The protein levels of phospho-ERK and ERK were detected by Western blotting (n=3). (B, D) Phospho-ERK protein concentrations were quantified by using Image J software and normalized to β-actin. Data are shown as the mean±standard, * P<0.05, ** P<0.01. In Press
11 Mar 2024 : Clinical Research
Comparison of Effects of Sugammadex and Neostigmine on Postoperative Neuromuscular Blockade Recovery in Pat...Med Sci Monit In Press; DOI: 10.12659/MSM.942773
12 Mar 2024 : Clinical Research
Comparing Neuromuscular Blockade Measurement Between Upper Arm (TOF Cuff®) and Eyelid (TOF Scan®) Using Miv...Med Sci Monit In Press; DOI: 10.12659/MSM.943630
11 Mar 2024 : Clinical Research
Enhancement of Frozen-Thawed Human Sperm Quality with Zinc as a Cryoprotective AdditiveMed Sci Monit In Press; DOI: 10.12659/MSM.942946
12 Mar 2024 : Database Analysis
Risk Factors of Age-Related Macular Degeneration in a Population-Based Study: Results from SHIP-TREND-1 (St...Med Sci Monit In Press; DOI: 10.12659/MSM.943140
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952