16 March 2022: Animal Study
A New Target of the Four-Herb Chinese Medicine for Wound Repair Promoted by Mitochondrial Metabolism Using Protein Acetylation Analysis
Zhongzhi Zhou1ABCDEFG, Li Chen23ABCDEFG, Yuxin Su4ABCDEF, Meirong Li3CE, Lingzhi Zhong3BCD, Liangying Liao1DEF, Yarong Ding2CEF, Mulin Liu2CDE, Zhonghang Yuan2CDE, Wei Wang2CEF, Xinyu Hang2BC, Xiuna Fang2DE, Xiaobing Fu3AEG*, Wenjun He3ADEF, Qian Hou3ABFGDOI: 10.12659/MSM.934816
Med Sci Monit 2022; 28:e934816






Abstract
BACKGROUND: Wound healing is a dynamic and complex process that is regulated by a variety of factors and pathways. This study sought to identify the mechanisms of the four-herb Chinese medicine ANBP in enhancing wound repair.
MATERIAL AND METHODS: By comparing the group treated with ANBP for 6 h (Z6h) with the corresponding control group (C6h), we used the new high-throughput differential acetylation proteomics method to explore the mechanism of ANBP treatment and analyse and identify new targets of ANBP for promoting wound healing.
RESULTS: ANBP promoted skin wound healing in mice; the wound healing process was accelerated and the wound healing time was shortened (P<0.05). The upregulated proteins were distributed mostly in the mitochondria to nuclear respiratory chain complexes and cytoplasmic vesicles. The dominant pathways for upregulated proteins were fatty acid metabolism, pyruvate metabolism, and tricarboxylic acid cycle. Pdha1 was upregulated with the most acetylation sites, while the downregulated Ncl, and Pfkm were most acetylated.
CONCLUSIONS: The findings from our study showed that ANBP improved cell aerobic respiration through enhanced glycolysis, pyruvic acid oxidative decarboxylation, and the Krebs cycle to produce more ATP for energy consumption, thus accelerating wound repair of skin.
Keywords: Acetylation, herbal medicine, Proteomics, Wound Healing, Animals, Cytokines, Disease Models, Animal, Male, Medicine, Chinese Traditional, Mice, Mitochondria, Signal Transduction, Skin, Wounds and Injuries
Background
Wound healing is a dynamic and complicated process regulated by many factors [1]. Once the protective barrier formed by the epidermis and dermis breaks down, the physiological process of wound healing takes place immediately [2]. Wound healing is composed of some overlapping phases involving clotting, inflammation, proliferation, and remodeling. During these processes, a series of complex biochemical events occur in a tightly coordinated cascade to repair the damage [3].
Recently, traditional Chinese medicine (TCM) has been increasingly recognized worldwide and extensively applied in the prevention and treatment of various diseases, such as cancer, diabetes mellitus, and skin disorders [4]. There is a rich experience with the use of TCM in trauma treatment in China’s long-term medical practice [5,6], with advantages of high efficacy, safety, and convenience [7]. Thefour-herb Chinese medicine ANBP is composed of
Material and Methods
PLANT MATERIAL:
All materials of ANBP were purchased from the First Affiliated Hospital of Hunan University of Chinese Medicine (Changsha, China) and were authenticated by the authors. The ANBP powder was deposited at Hunan University of Chinese Medicine (Changsha, China). The batch numbers were: Agrimonia eupatoria L. (Hubei, China), Nelumbon ucifera Gaertn (Mongolia, China), Boswellia carteri Biedw. (Hunan, China), and Typha orientalis C. Presl (Hunan, China).
STUDY DESIGN:
We purchased 24 male, 4-week-old, 18±2 g, SPF C57BL/6 mice from Hunan SJA Laboratory Animal Co. The housing conditions were: temperature 20~22°C, relative humidity 40~60%, 12 h of light, 12 h of light protection. The company was equipped with quarantine facilities, and all of the animals were regularly quarantined without pathogenic microorganisms.
Twenty-four mice were randomly divided into a control group and a treatment group. After intraperitoneal injection with 10% hydrated chloral aldehyde (6 mg/kg), hair was removed from the dorsal skin of the mice using depilation cream. A 1-cm-diameter circle was marked at the depilation site of the back and a cut was made down along the mark through the entire skin layer to the muscle layer, resulting in the full-layer skin defect model of the mouse. In the treatment group, the wound area was immediately covered with ANBP powder (each mouse received ANBP 90 mg each time, including 30 mg of
EXTRACTING PROTEIN:
First, the sample was ground in liquid nitrogen. After that, the powder was placed into a 5-mL centrifuge tube and dissociated on ice 3 times using an ultrasonic instrument in cold lysis buffer (1% protease inhibitor cocktail, 2 mM EDTA, 8 M urea, and 10 mM DTT). Then, the supernatant was delivered to new tubes for centrifugation at 4°C at 20 000 g for 10 min, and the remaining debris was discarded. BCA kit (Beyotime Biotechnology, China) was used to detect protein concentrations.
TRYPSIN DIGESTION:
The proteins were reduced with 10 mM DTT (Sigma-Aldrich, USA) at 37°C for 60 min and alkylated using 20 mM IAA at room temperature in darkness environment for 45 min. For trypsin digestion, 100 mM TEAB (Sigma-Aldrich, USA) was added to the samples until the urea concentration was less than 2 M. Finally, adding trypsin for the first digestion overnight at a mass ratio of trypsin-to-protein of 1: 50; the second digestion at a mass ratio of trypsin-to-protein of 1: 100 for 4 h. For each sample, approximately, 100 μg of protein was digested using trypsin in the following experiments.
LABELING WITH TMT:
A Strata X C18 SPE column (Phenomenex, USA) was used to desalt the peptide after trypsin digestion, and then the peptide was dried in a vacuum. After the peptide reconstituted in 0.5 M TEAB, it was then processed for the TMT kit (ThermoFisher Scientific, USA) in accordance with the manufacturer’s protocol. Briefly, 1 unit (defined as the amount of reagent required to label 100 μg of protein) of TMT reagent was thawed and reconstituted in 24 μL of ACN. The peptide mixtures were incubated at room temperature for 2 h, and then were desalinated and dehydrated by vacuum centrifugation.
FRACTIONATION WITH HPLC:
Using a Thermo Betasil C18 column (5 μm particles, 10 mm ID, 250 mm length), high-pH reverse-phase HPLC (Agilent, USA) was used to fractionate the protein samples. Briefly, with 8% to 32% gradient acetonitrile, peptides were first separated in 10 mM ammonium bicarbonate at pH 10 into 60 fractions after 60 min. After that, the peptides were combined into 6 fractions and finally dried by centrifuging in a vacuum.
AFFINITY ENRICHING ACETYLATED PEPTIDES:
For enriching acetylated peptides, NETN buffer (1 mM EDTA, 50 mM Tris-HCl, 100 mM NaCl, 0.5% NP-40, pH 8.0) was used to dissolve the tryptic peptides, and then the peptides and the prewashed antibody beads (PTM Biolabs, China) were incubated overnight at 4°C. The beads were washed with NETN buffer and ddH2O. The bound peptides were eluted from the beads with 0.1% TFA. Elution peptides were combined and dried in a vacuum. The resulting peptides were washed with C18 ZipTips (Merck Millipore, USA), followed by analysis with LC-MS/MS.
ANALYSIS WITH LC-MS/MS:
Peptides were first dissolved in 0.1% FA for proteome analysis and immediately transferred into a precolumn (Acclaim PepMap RSLC, ThermoFisher Scientific, USA) in reversed phase. The gradient consisted of a progressive increase from 8% to 22% solution B (0.1% FA in 98% ACN) for 19 min, 22% to 35% for 13 min, then increased to 80% for 4 min, and was maintained, all at a steady flow rate of 0.8 μL/min in an EASY-nLC 1000 UPLC system.
Briefly, the peptide was exposed to an NSI source, then underwent tandem mass spectrometry (MS/MS) in a Q Exactive™ Plus (ThermoFisher Scientific, USA) connected online to the UPLC. The intact peptide was determined in the Orbitrap at a resolution of 70 000. Using the NCE setting of 35, the peptide was selected for MS/MS; ion fragments were tested at a resolution of 17 500 in the Orbitrap. A data-dependent analysis process alternating between 1 MS scan and 10 MS/MS scans was utilized for the top 20 precursor ions beyond the 1E4 ion count threshold in the MS survey scan with 30% dynamic exclusion. The electrospray voltage utilized was 2000 V. Overfilling of the ion trap was prevented by using automatic gain control (AGC); MS/MS spectra were generated by accumulating 5E4 ions. The m/z scan range was 350 to 1800 for MS scans, and the fixed first mass was set up as 100 m/z.
SEARCHING THE DATABASE:
The resultant MS/MS data were analyzed using MaxQuant installed with the Andromeda search engine (v.1.5.2.8). The SwissProt
QUALITY CONTROL VALIDATION OF MS DATA:
We used the mass error distribution of all identified peptides and peptide length distribution to make the quality control of MS data validation. First, the mass error of all the identified peptides were examined. The mass error distribution was near zero, and most were <0.02 Da. This result indicates that the MS data mass accuracy meets the requirement. Second, most peptide lengths were distributed from 8 to 20, which conforms to the tryptic peptide property, meaning that the preparation of samples reached the criterion.
BIOINFORMATICS ANNOTATION FOR DIFFERENTIAL PROTEINS:
The Gene Ontology (GO) annotation proteome was provided by the UniProt-GOA database (
The protein pathway was noted by using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. First, the KEGG online service tool KAAS was used to depict the description of the proteins’ KEGG database. Second, we used the KEGG online service tool KEGG mapper to map the annotation results in the KEGG pathway database. The enrichment of differentially expressed proteins against all identified proteins was tested by using a two-tailed Fisher’s exact test. A corrected
FUNCTIONAL ENRICHMENT OF DIFFERENTIAL PROTEINS:
Via GO annotation, proteins were divided into 3 categories: molecular function, cellular compartment, and biological process. For each of these categories, the enrichment of diverse expressed proteins relative to all identified proteins was determined by two-tailed Fisher’s exact test. Using standard false discovery rate control methods, a large amount of hypothesis testing was corrected. GO terms with
The enriched pathways were identified using the KEGG database, and the enrichment of dissimilar expressed proteins against all identified proteins was tested by two-tailed Fisher’s exact test. Standard false discovery rate control methods were used to correct for multiple hypothesis testing. Pathways with
MOTIF AND STATISTICAL ANALYSIS:
Data analysis was performed using SPSS 25.0. For multiple groups of measurement data, the samples were tested for normal distribution and homogeneity of variance. If they met the conditions of normality, one-way ANOVA was used. When the variance was homogeneous, LSD was used for multiple comparison, and games Howell was used for multiple comparison when the variance was uneven.
The amino acid sequence model in particular positions of modify-21-mers (10 amino acids downstream and upstream of the sites) in all sequences of proteins was calculated using soft Motif-X. We used all protein sequences in the database for background database parameters, and other parameters were default values.
Results
ANBP ACCELERATES WOUND HEALING IN MOUSE SKIN:
We confirmed that ANBP could promote skin wound healing in mice. At 6 h, 3 days, 7 days, and 14 days after wounding, the images of the wound were recorded, and the wound closure of each group was calculated at each time point. Compared with the control group, ANBP treatment significantly reduced bleeding and edema, and accelerated wound healing in mouse skin (Figure 1A). The statistical results showed that the wound healing rate of ANBP group at 3 days, 7 days, and 14 days was significantly higher than that of the control group, and the difference was statistically significant (Figure 1B).
GENERATION OF THE GLOBAL PROTEOME AND ACETYLATED PROTEOME:
To comprehensively analyze the protein expression in skin tissues from the ANBP treatment group and the control group, we used an up-to-date proteomic strategy to scan the proteome and acetylated proteome. Based on an integrated approach involving TMT labeling, protein prefractionation by HPLC, and specific affinity enrichment for acetylated peptides and then LC-MS/MS, followed by bioinformatics analysis (Figure 2), we generated the global proteome and acetylated proteome, providing data illustrating the mechanism of wound healing promoted by ANBP. In detail, the whole experimental process included 4 main steps: (a) establishment of the mouse model, drug treatment, photographing, and collection of samples; (b) protein extraction, trypsin digestion, HPLC score and iTRAQ labeling; (c) analysis of LC-MS/MS; and (d) bioinformatics analysis.
QUALITY CONTROL VALIDATION OF MS DATA:
The mass errors of all the identified peptides were checked to verify the quality of MS and MS/MS profiling (Supplementary Figure 1). The peptide mass error distribution was close to zero, and most were <2 ppm, indicating that the data of MS mass accuracy met the requirements. Most peptides were distributed between 7 and 16 in length, which conformed to the tryptic peptide properties, demonstrating that the sample preparation fit the criterion.
To evaluate the repeatability of the 3 biological replications, the Pearson correlation coefficient was used to perform the repeatability analysis. For the global proteome and acetylated proteome profiling shown in Supplementary Figure 2, the Pearson correlation coefficients of biological samples in triplicate were all >0.8, but the correlation between different groups was poor, demonstrating that each group of the experiment had good biological repeatability. Therefore, the proteomic analysis was reliable, and all data were available for subsequent high-quality analysis.
DATA FOR THE GLOBAL PROTEOME AND ACETYLATED PROTEOME:
Based on the above strategy and data quality control, we then achieved comprehensive protein profiling with identification and quantification, as shown in Table 1. A total of 3238 proteins were identified, with 2581 quantified proteins in the global proteome (Supplemental Table 1), 485 identified acetyl proteins with 1032 acetylation sites, and 449 quantified acetyl proteins with 948 acetylation sites in the acetylated proteome (Supplementary Table 2).
COMPARISON OF DIFFERENTIALLY QUANTIFIED PROTEINS:
At 6 h of early wound healing, the expression of acetylated modified proteins in the skin of the ANBP treatment group (Z6h) was compared with that in the control group (C6h). The amounts of the differentially expressed proteins are summarized in Table 2 by the difference with an average cut-off change of 1.2-fold and P<0.05. There were 101 upregulated proteins and 282 downregulated proteins in the global proteome (Supplementary Table 3). For the acetylome, 86 acetyl proteins were upregulated with 110 acetyl sites, and 31 were downregulated with 45 acetyl sites (Supplementary Table 4).
After ANBP treatment, the Motif-X tool was used to extract the motifs of the identified acetylated peptides (10 downstream and 10 upstream amino acids of the acetylated lysine site) to confirm the lysine acetylation preferences for significantly altered acetylation sites. We matched amino acids at the background and foreground of the acetylation site from the whole size and calculated motif scores and fold changes; thus, we could speculate on the possible amino acids near the acetylated site and obtain statistical significance. We obtained the 7 most likely sequences around the acetylated site, where KK, KH, KN, and KR exhibited the highest motif scores, as shown in Table 3. Lysine was still most active near the acetylated site.
To explain the sequence characteristics’ quantitative diversity around the specific acetylation sites after ANBP treatment, we showed motif enrichment for downstream and upstream amino acids of acetylation sites as a heatmap (Figure 3). Arginine, asparagine, lysine, and histidine were preferentially situated at the +1 position, and lysine was also enriched at the +4 and +5 positions. In addition, glycine was preferentially located at the −1 position, followed by alanine or aspartic acid, and, surprisingly, histidine was specifically enriched at the −2, −3, and −4 positions, while arginine was enriched at the −6 and −7 positions. Evidently, the +1 position is more variable and is the preferred site flanking the acetylated lysine.
FUNCTIONAL ANNOTATION FOR DIFFERENTIALLY QUANTIFIED ACETYL SITES AFTER ANBP TREATMENT:
To reveal the function and subcellular localization of differentially acetylated proteins after ANBP treatment, bioinformatics analysis and subcellular function annotation were conducted based on Gene Ontology (GO), including molecular function, biological process, and cellular component analyses. For the biological process classification in GO annotation, the largest group was cellular process, followed by single-organism process, metabolic process, and biological control (Table 4). The first 3 cell components were organelle, cell, and extracellular regions. Changes in molecular function included binding and catalytic activity, making up the majority of differentially acetylated proteins. The differential expression protein was mainly located in the cytoplasm (53% and 34%), mitochondria (10% and 33%), and nucleus (20% and 19%) for the upregulated and downregulated proteins, respectively (Figure 4).
CLUSTERING ANALYSIS OF ACETYLATION SITES IN WOUNDED SKIN TREATED WITH ANBP:
To investigate the properties of quantified proteins after ANBP treatment in wounded skin, clustering analysis was performed based on GO enrichment. As shown in Figure 5A, all differentially acetylated sites were divided into 4 groups on the basis of the quantitative ratios Q1 (ratio <0.76), Q2 (0.76< ratio <0.83), Q3 (1.2< ratio <1.3), and Q4 (ratio >1.3). The number of differential acetylated sites was 31 in Q1, 14 in Q2, 47 in Q3, and 63 in Q4. Based on the enrichment of clustering analysis, the upregulated proteins were involved in many processes at both Q3 and Q4, including cell respiration, energy production by lipid oxidation, tricarboxylic acid metabolism, redox process, the Krebs cycle, citrate metabolism, acetyl CoA metabolism and metabolites and energy metabolism precursors and aerobic respiration, while metabolic processes of coenzyme, cofactor, and carbohydrate were enriched in the downregulated proteins (Figure 5B). For the cellular component, clustering analysis showed that the upregulated proteins were distributed mostly in the mitochondria to the nucleus, respiratory chain complexes, and cytoplasmic vesicles, while proteins associated with extracellular vesicles, microtubules, and ribonucleoprotein granules were downregulated (Figure 5C). Based on molecular function enrichment, clustering analysis showed an increase in protein expression in binding activity, dehydrogenase activity, and carbon-oxygen lyase activity, as well as a decrease in kinase regulator activity, kinase inhibitor activity, and nucleic acid binding (Figure 5D).
To identify the pathways involved in ANBP treatment, KEGG pathway clustering analysis and protein domain analysis were further carried out (Figure 6). Based on 4 groups of all differentially acetylated proteins (Q1, Q2, Q3, and Q4), the dominant pathways for upregulated proteins were fatty acid metabolism, pyruvate metabolism, and tricarboxylic acid cycle, while the downregulated proteins included vascular smooth muscle contraction, regulation of actin cytoskeleton, and glutathione metabolism (Figure 6A). For clustering analysis with the protein structure domain, the upregulated proteins were involved in the pleckstrin homology domain, disulfide isomerase, thioredoxin, and thiolase-like domain after ANBP treatment, while the downregulated proteins were mainly distributed in globulin, globin-like, and myosin (Figure 6B).
PROTEIN INTERACTION NETWORK OF DIFFERENTIALLY ACETYLATED SITES FOR ANBP TREATMENT:
When skin tissues were treated with ANBP, a protein-protein interaction network of acetyl groups was established. We analysed the protein-protein interaction network based on the acetylated proteins to identify the main nodes and important connections after periposine treatment. There were 79 nodes and 418 connections seen from the constructed protein-protein interaction network. Ttn, Suclg1, Uqcrc2, Myl1, Tnnt3, Actn3, Tnni2, Dld and EP300 were the top 8 centre nodes. As illustrated from the network (Figure 7), Pdha1 was upregulated with the most acetylation sites, while the downregulated Ncl, and Pfkm were most acetylated. In addition, Hist1h1e, Myh4 and Hist1h2bc had both upregulated and downregulated acetylation sites, which may participate in divergent functions.
Discussion
The four-herb Chinese medicine ANBP is mainly used for the early treatment of extensive trauma [12]. Compared with the control group, ANBP treatment not only significantly enhanced wound healing, but also reduced scar formation in our previous research [20]. Our further comparative proteomic analysis indicated that ANBP significantly promoted angiogenesis-related process and many hemostasis/coagulation regulation-related processes, and demonstrated that proteins differentially expressed in the micronized ANBP group were mainly localized in the mitochondria in the early healing phase [8].
When skin, connective tissue, and muscle are damaged in a wound, cell mitochondria will also experience structural destruction and dysfunction [21]. Mitochondrial structure damage and functional transformation are key factors during the wound healing process [22,23]. The inner membrane, as well as the membrane space between the inner and outer membranes, contains various enzymes closely related to ATP synthesis, which are also directly involved in a variety of metabolic pathways [24]. At present, more than 400 posttranslational protein modifications have been reported, with the deepest and widest understanding including glycosylation, phosphorylation, and acyl modifications, such as methylation, acetylation, ubiquitin, and succinyl modification [25], which play important roles in controlling protein structure, function, and activity [26]. Among them, succinylation and acetylation play significant roles in signal transmission, mitochondrial respiration, inflammatory immunity, cell regulation, metabolic regulation, and other physiological processes and diseases [27].
Proteomics research of full-thickness skin defects in mouse backs at 6 h, 3 days, 7 days, and 14 days showed that differential protein expression was mainly involved in the early wound healing of ANBP intervention, and the subcellular localization was enriched in mitochondria. In this study, ANBP enhanced wound healing in the dorsal skin of mice, and quantitative acetylation proteomics analysis of mouse skin at 6 h after wound showed that glycolysis was enhanced (Figure 1). NAD+ binding activity was increased, indicating NAD+ hydrogen binding ability was enhanced to provide a substrate for oxide reduction with the mitochondrial respiratory chain, promoting aerobic respiration and producing more energy to meet the high demand for energy during wound repair. Second, the oxidation and decarboxylation of pyruvate was promoted. Clustering of cellular components showed an obvious increase in the pyruvate dehydrogenase complex, similar to the molecular functions of acetyl transfer activity, NAD+ activity, NADP+ activity, and pyruvate dehydrogenase activity, as well as the biological process of acetyl CoA metabolism. KEGG pathway enrichment and clustering both revealed that pyruvate metabolism was significantly increased (Figure 6). Last, the Krebs cycle was accelerated. Substrates such as acetyl CoA, ketone glutaric acid, isocitrate dehydrogenase, ketone glutaric acid dehydrogenase complex, sulfur lipid hydrolase, and others were all increased in molecular function, cellular component, biological process, and KEGG pathway analyses. Importantly, the a-ketone glutaric acid dehydrogenase complex and malate dehydrogenase are key participants in NADH+H+ reduction in the Krebs cycle. The organelle capsule, mitochondrial membrane and mitochondrial respiratory chain were obviously enriched and increased in clustering. In addition, cellular component analysis showed high expression of mitochondrial nucleoids, respiratory chain complexes, and the highest upregulated tricarboxylic acid pathway through KEGG enrichment and clustering (Figure 6).
Only protein acetylation analysis was done, showing that ANBP improved cell aerobic respiration by promoting glycolysis, pyruvic acid oxidative decarboxylation, and the Krebs cycle to produce more ATP to speed wound repair of skin. However, the mechanism by which the four-herb Chinese medicine ANBP regulates related activities to promote wound healing requires further study.
Conclusions
In summary, the above results showed that ANBP improved cell aerobic respiration through enhanced glycolysis, pyruvic acid oxidative decarboxylation, and the Krebs cycle to produce more ATP for energy consumption, thus accelerating wound repair of skin. Therefore, in mitochondria as a target, understanding mitochondrial changes after trauma, as well as the effect on organisms and the underlying mechanism, provides a new idea and breakthrough to accelerate wound healing and improve the quality of wound healing as a new direction for the clinical treatment of trauma. The interaction of mitochondria with different types of trauma and the body’s basic situation vary under certain circumstances. As a result, although mitochondria are located at a core position in the process of wound healing, there should be a focus on how mitochondria have the potential to promote healing in different situations.
Figures
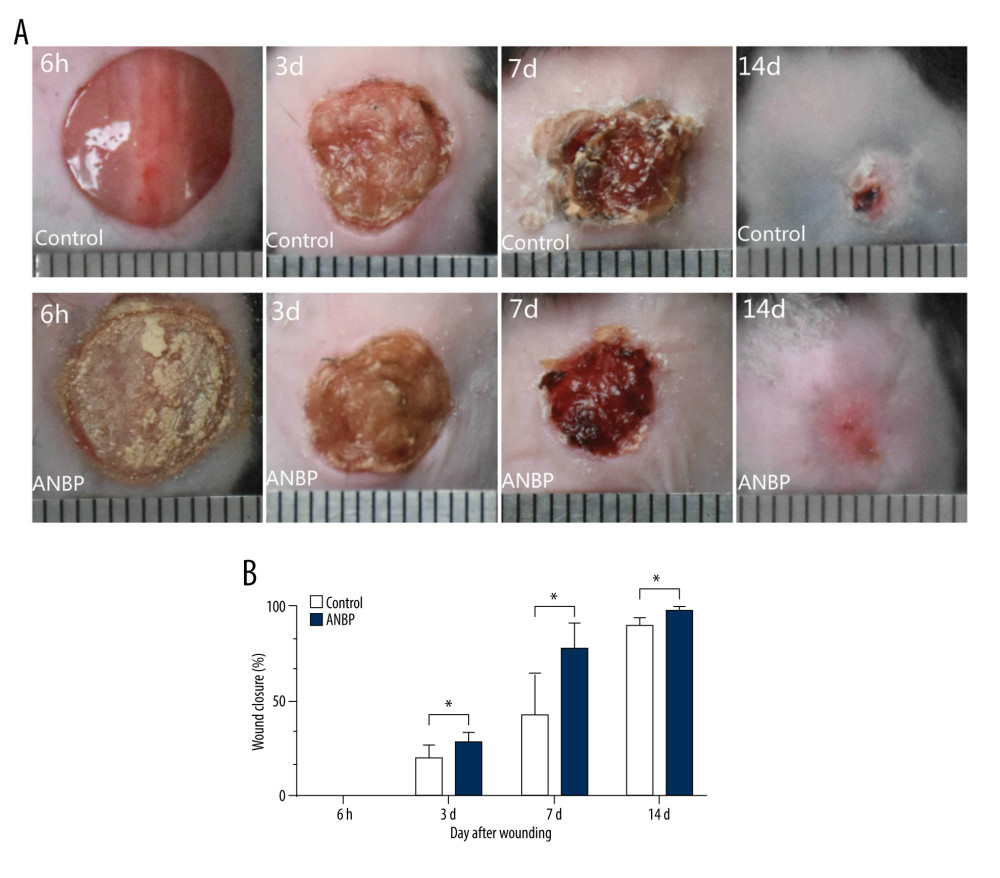 Figure 1. Effect of ANBP on wound healing. (A) Skin wound healing of mice in the ANBP treatment group and the control group at 6 h, 3 d, 7 d and 14 d after wounding. (B) Wound healing rate in the ANBP treatment group and the control group. The wound healing rate of the ANBP treatment group and the control group. Percentage wound healing = (initial wound size – current wound size) ÷ initial wound size ×100%.
Figure 1. Effect of ANBP on wound healing. (A) Skin wound healing of mice in the ANBP treatment group and the control group at 6 h, 3 d, 7 d and 14 d after wounding. (B) Wound healing rate in the ANBP treatment group and the control group. The wound healing rate of the ANBP treatment group and the control group. Percentage wound healing = (initial wound size – current wound size) ÷ initial wound size ×100%. 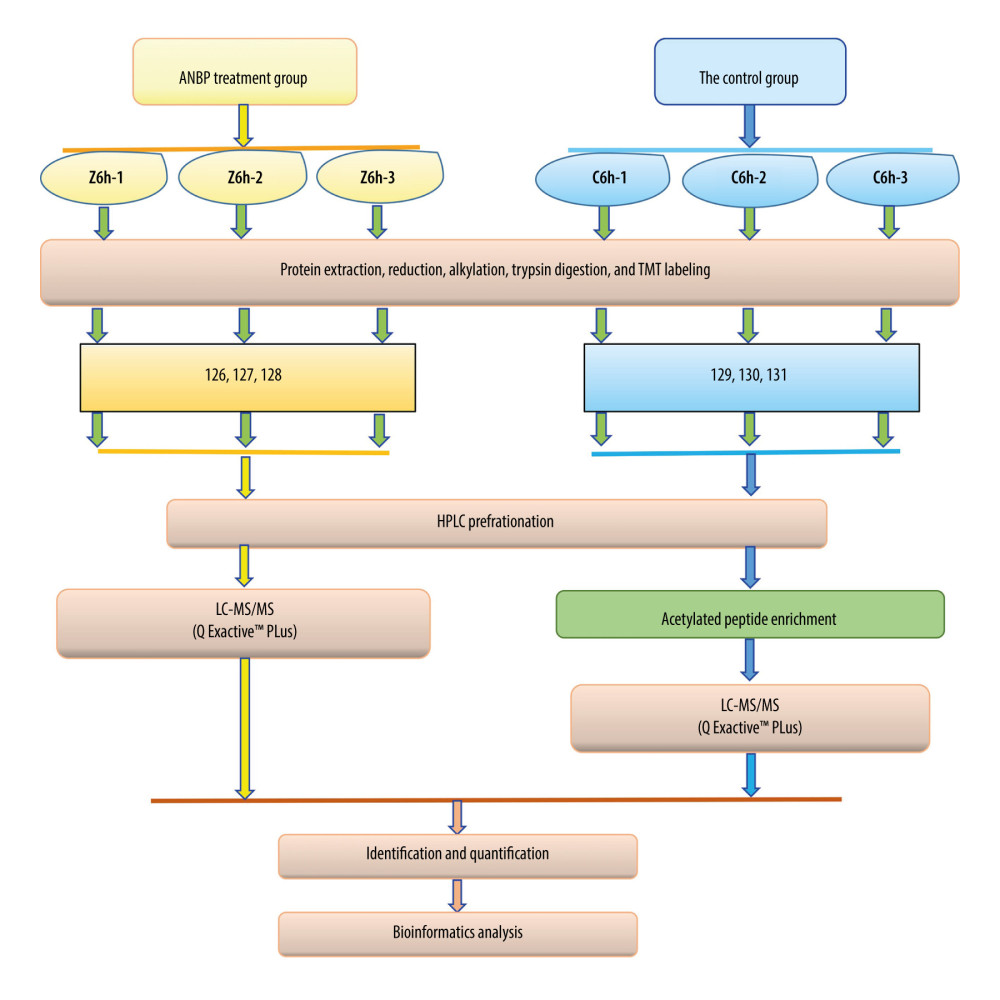 Figure 2. Workflow for the quantitative acetylome and proteome based on TMT. Three replicates of the global proteome and acetylated proteome were performed in the ANBP treatment group and the control group.
Figure 2. Workflow for the quantitative acetylome and proteome based on TMT. Three replicates of the global proteome and acetylated proteome were performed in the ANBP treatment group and the control group. 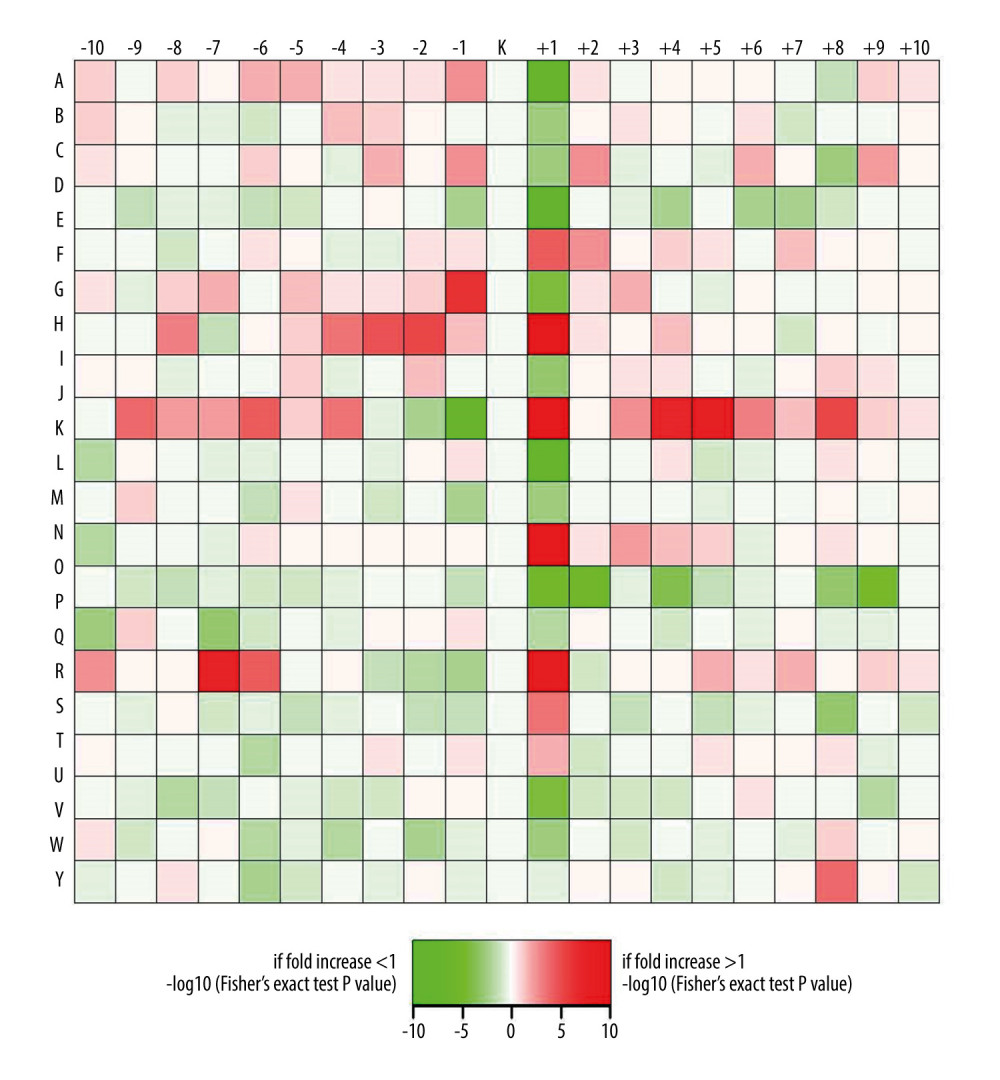 Figure 3. Lysine acetylation preferences for significantly changed acetylation sites. Red represents a significant concentration of this amino acid close to the modification site, while green represents a significant reduction in secondary amino acids close to the modification site.
Figure 3. Lysine acetylation preferences for significantly changed acetylation sites. Red represents a significant concentration of this amino acid close to the modification site, while green represents a significant reduction in secondary amino acids close to the modification site. 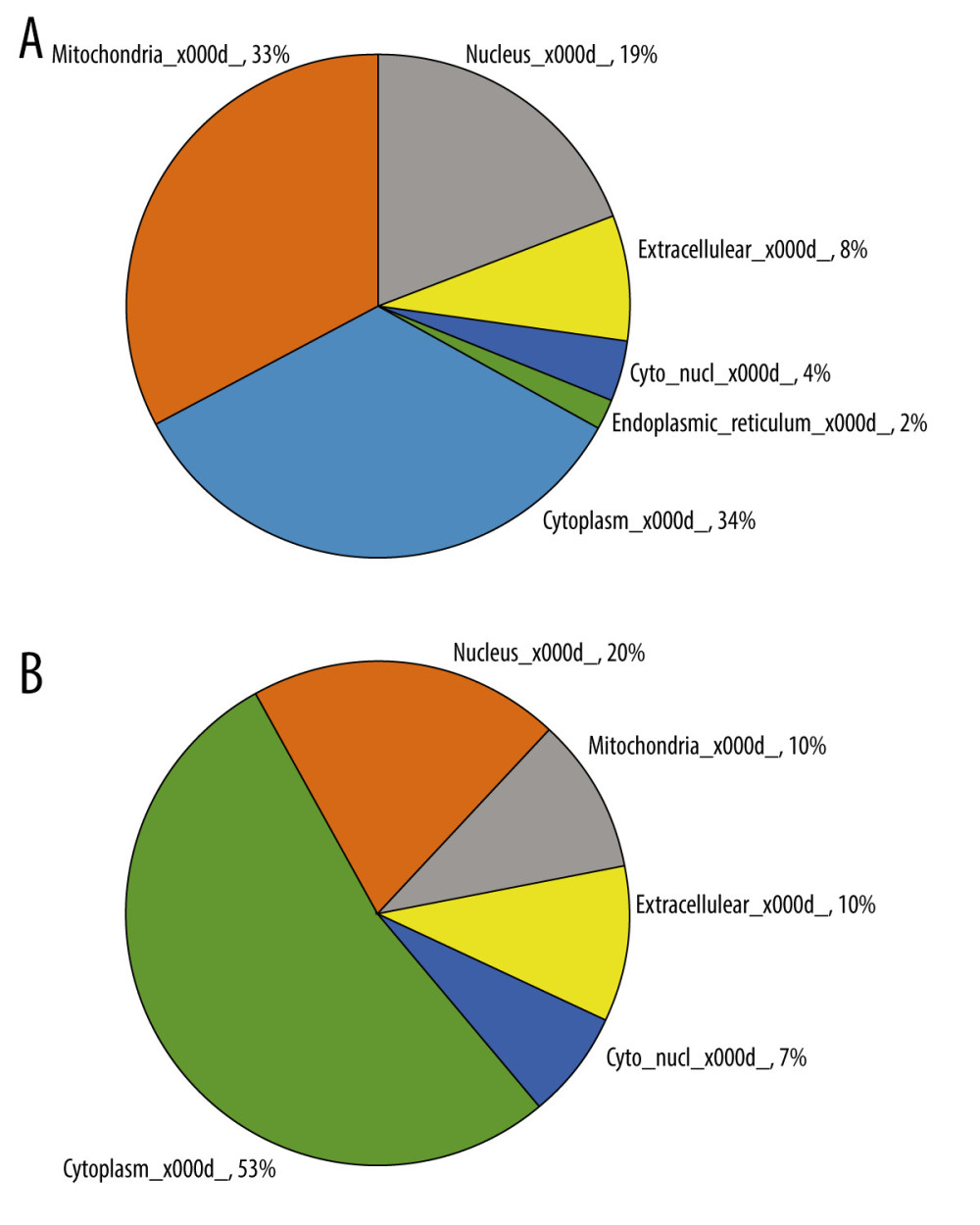 Figure 4. Distribution of subcellular location for upregulated and downregulated proteins. (A) Upregulated protein subcellular location in Z6h/C6h; (B) downregulated protein subcellular location in Z6h/C6h.
Figure 4. Distribution of subcellular location for upregulated and downregulated proteins. (A) Upregulated protein subcellular location in Z6h/C6h; (B) downregulated protein subcellular location in Z6h/C6h.  Figure 5. Classification of differentially acetylated sites and heatmap of clustering analysis based on GO enrichment. (A) Classification of differentially acetylated sites Q1, Q2, Q3 and Q4. (B) Heatmap of biological processes for the 4 groups of differentially acetylated sites. (C) Heatmap of cellular components for the 4 groups of differentially acetylated sites. (D) Heatmap of molecular functions for the 4 groups of differentially acetylated sites.
Figure 5. Classification of differentially acetylated sites and heatmap of clustering analysis based on GO enrichment. (A) Classification of differentially acetylated sites Q1, Q2, Q3 and Q4. (B) Heatmap of biological processes for the 4 groups of differentially acetylated sites. (C) Heatmap of cellular components for the 4 groups of differentially acetylated sites. (D) Heatmap of molecular functions for the 4 groups of differentially acetylated sites. 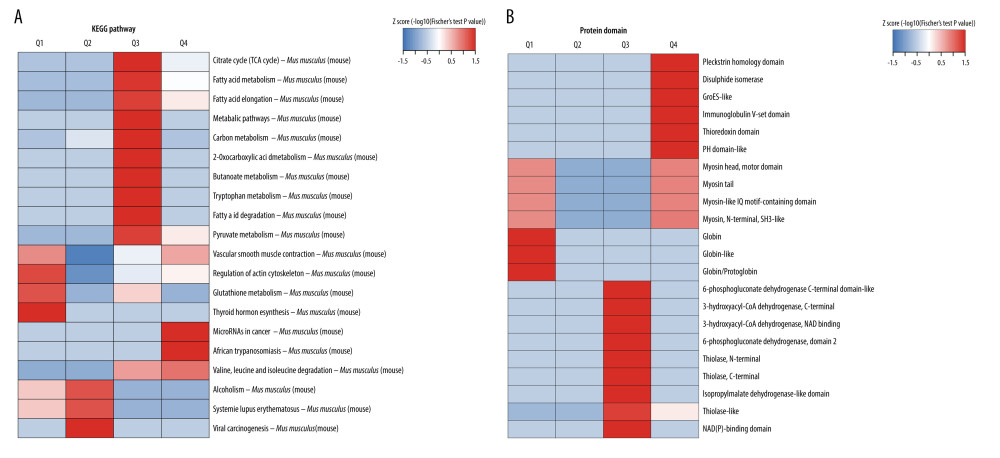 Figure 6. Heatmap of clustering analysis based on KEGG pathways and protein domains. (A) Heatmap of KEGG pathways for the 4 groups of differentially acetylated sites. (B) Heatmap of protein domains for the 4 groups of differentially acetylated sites.
Figure 6. Heatmap of clustering analysis based on KEGG pathways and protein domains. (A) Heatmap of KEGG pathways for the 4 groups of differentially acetylated sites. (B) Heatmap of protein domains for the 4 groups of differentially acetylated sites. 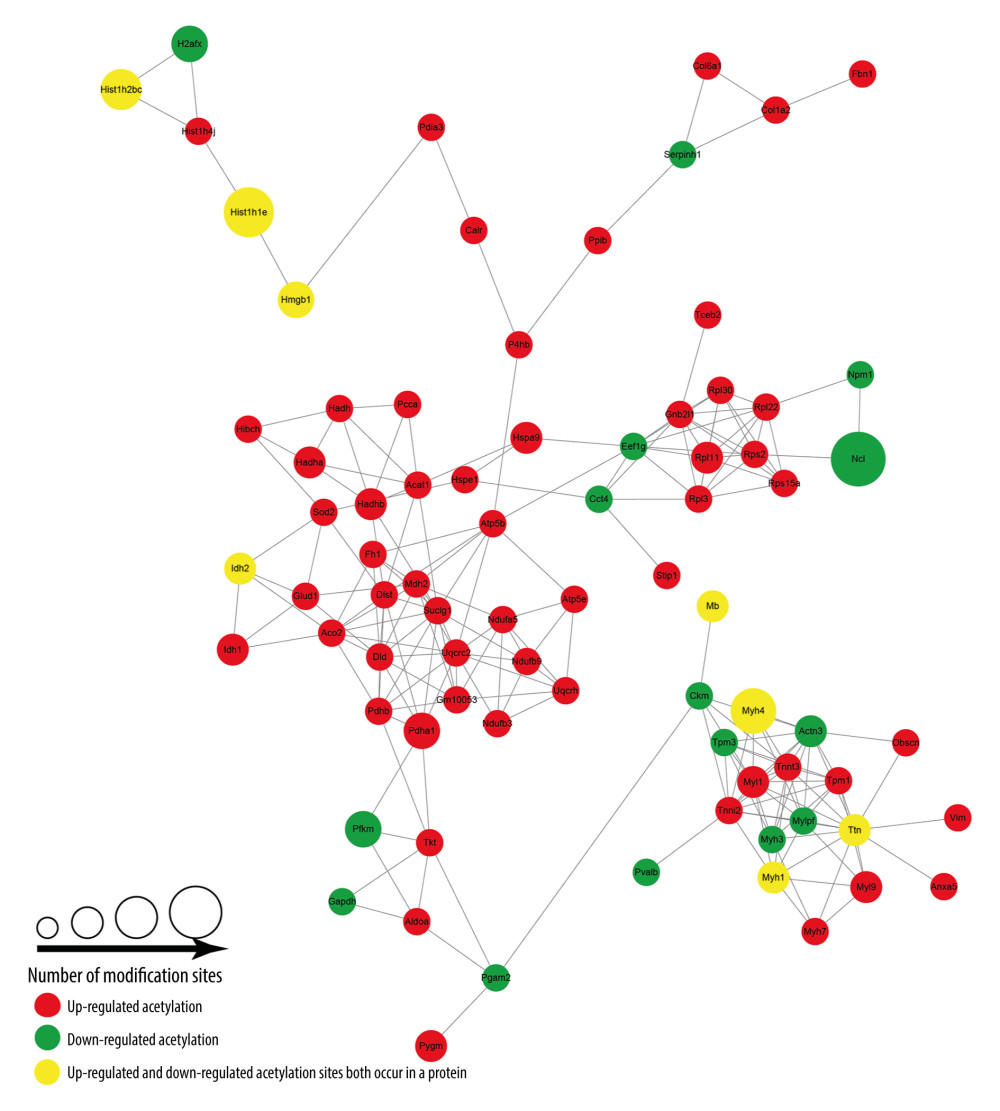 Figure 7. Protein–protein interaction network of acetylated proteins in periposine-treated AS cells.
Figure 7. Protein–protein interaction network of acetylated proteins in periposine-treated AS cells. Tables
Table 1. Comprehensive profiling of the global and acetylated proteomes.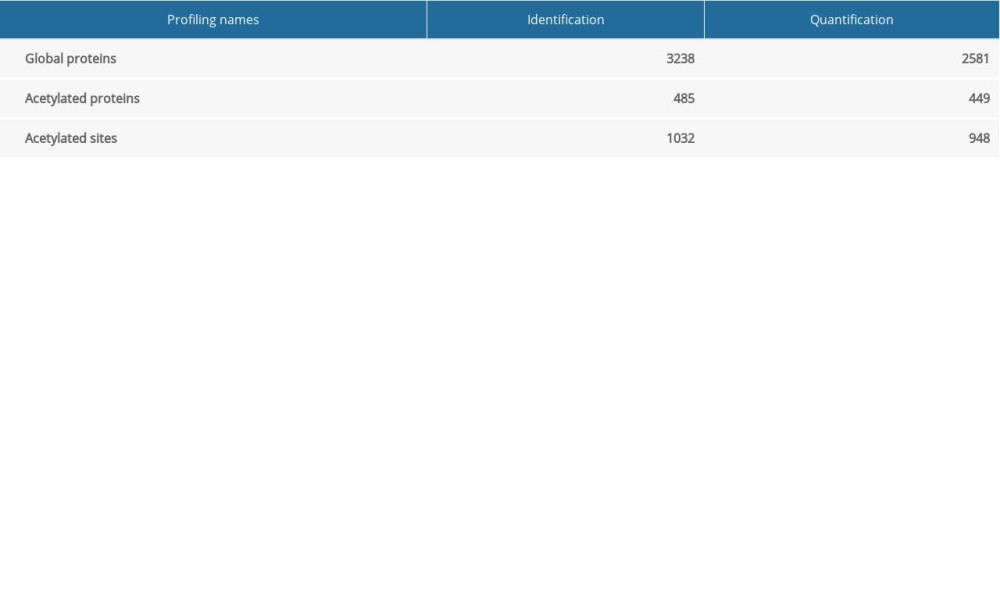 Table 2. Number of differentially quantified proteins by comparing ANBP-treated mice with the control.
Table 2. Number of differentially quantified proteins by comparing ANBP-treated mice with the control. Table 3. The feature sequence around acetylated sites by motif analysis.
Table 3. The feature sequence around acetylated sites by motif analysis.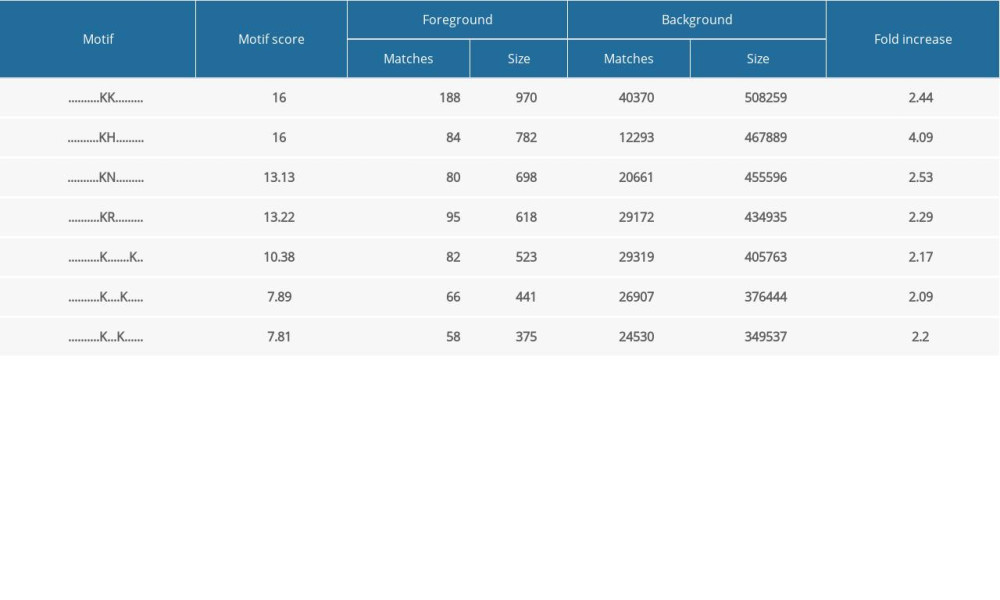 Table 4. GO annotation for upregulated and downregulated acetylated proteins.
Table 4. GO annotation for upregulated and downregulated acetylated proteins.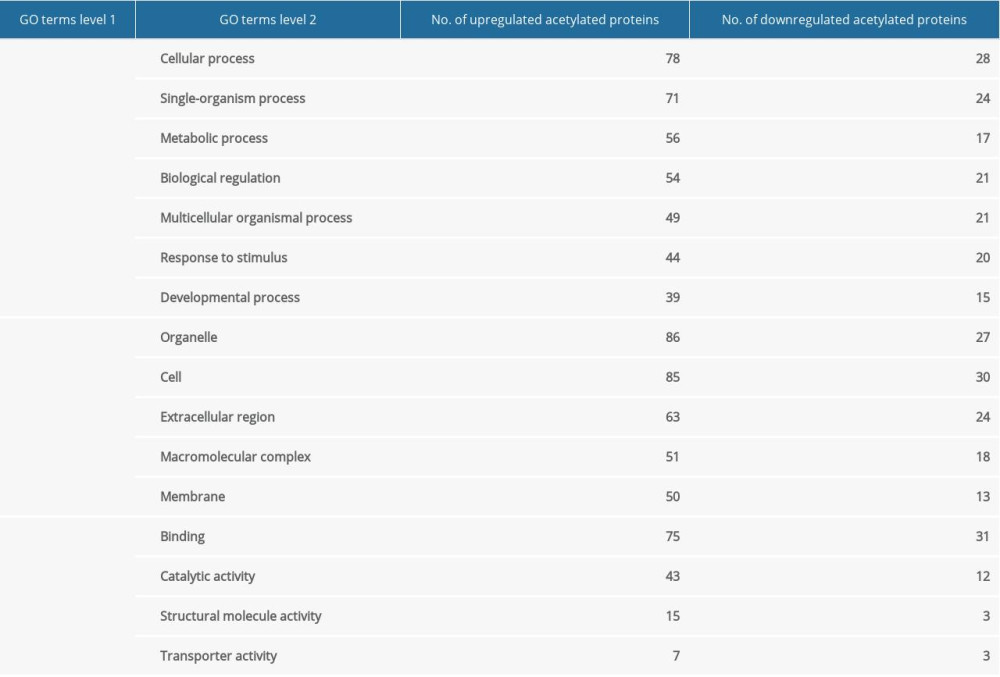
References
1. Takeo M, Lee W, Ito M, Wound healing and skin regeneration: Cold Spring Harb Perspect Med, 2015; 5; a023267
2. Kasuya A, Tokura Y, Attempts to accelerate wound healing: J Dermatol Sci, 2014; 76; 169-72
3. Brown A, Phases of the wound healing process: Nurs Times, 2015; 111; 12-13
4. Qi F, Zhao L, Zhou A, The advantages of using traditional Chinese medicine as an adjunctive therapy in the whole course of cancer treatment instead of only terminal stage of cancer: Biosci Trends, 2015; 9; 16-34
5. Leung PC, Ko EC, Siu WS, Selected topical agents used in traditional Chinese medicine in the treatment of minor injuries – a review: Front Pharmacol, 2016; 7; 16
6. Wang R, Lechtenberg M, Sendker J, Wound-healing plants from TCM: In vitro investigations on selected TCM plants and their influence on human dermal fibroblasts and keratinocytes: Fitoterapia, 2013; 84; 308-17
7. Chak KF, Hsiao CY, Chen TY, A study of the effect of Shiunko, a traditional Chinese herbal medicine, on fibroblasts and its implication on wound healing processes: Adv Wound Care (New Rochelle), 2013; 2; 448-55
8. Chen L, Hou Q, Zhou ZZ, Comparative proteomic analysis of the effect of the four-herb Chinese medicine ANBP on promoting mouse skin wound healing: Int J Low Extrem Wounds, 2017; 16; 154-62
9. Shen Y, Guan Y, Song X: Food Sci Nutr, 2019; 7; 3062-70
10. Gao M, Ge Z, Deng R, Evaluation of VEGF mediated pro-angiogenic and hemostatic effects and chemical marker investigation for Typhae Pollen and its processed product: J Ethnopharmacol, 2021; 268; 113591
11. Chen P, Liu S, Dai G, Determination of typhaneoside in rat plasma by liquid chromatography-tandem mass spectrometry: J Pharm Biomed Anal, 2012; 70; 636-39
12. Hou Q, He WJ, Chen L, Effects of the four-herb compound ANBP on wound healing promotion in diabetic mice: Int J Low Extrem Wounds, 2015; 14; 335-42
13. Cano Sanchez M, Lancel S, Boulanger E, Neviere R, Targeting oxidative stress and mitochondrial dysfunction in the treatment of impaired wound healing: A systematic review: Antioxidants (Basel), 2018; 7; 98
14. Schiffmann LM, Werthenbach JP, Heintges-Kleinhofer F, Mitochondrial respiration controls neoangiogenesis during wound healing and tumour growth: Nat Commun, 2020; 11; 3653
15. Beltrao P, Bork P, Krogan NJ, van Noort V, Evolution and functional cross-talk of protein post-translational modifications: Mol Syst Biol, 2013; 9; 714
16. Hofer A, Wenz T, Post-translational modification of mitochondria as a novel mode of regulation: Exp Gerontol, 2014; 56; 202-20
17. Sabari BR, Zhang D, Allis CD, Zhao Y, Metabolic regulation of gene expression through histone acylations: Nat Rev Mol Cell Biol, 2017; 18; 90-101
18. Herskovitz I, Hughes OB, Macquhae F, Epidermal skin grafting: Int Wound J, 2016; 13(Suppl 3); 52-56
19. Gorecka J, Kostiuk V, Fereydooni A, The potential and limitations of induced pluripotent stem cells to achieve wound healing: Stem Cell Res Ther, 2019; 10; 87
20. Hou Q, He WJ, Hao HJ, The four-herb Chinese medicine ANBP enhances wound healing and inhibits scar formation via bidirectional regulation of transformation growth factor pathway: PLoS One, 2014; 9; e112274
21. Lim TK, Rone MB, Lee S, Antel JP, Zhang J, Mitochondrial and bioenergetic dysfunction in trauma-induced painful peripheral neuropathy: Mol Pain, 2015; 11; 58
22. Janda J, Nfonsam V, Calienes F, Modulation of ROS levels in fibroblasts by altering mitochondria regulates the process of wound healing: Arch Dermatol Res, 2016; 308; 239-48
23. Papanicolaou KN, Rourke B, Foster DB, Metabolism leaves its mark on the powerhouse: Recent progress in post-translational modifications of lysine in mitochondria: Front Physiol, 2014; 5; 301
24. Qureshi MA, Haynes CM, Pellegrino MW, The mitochondrial unfolded protein response: Signaling from the powerhouse: J Biol Chem, 2017; 292; 13500-6
25. Lothrop AP, Torres MP, Fuchs SM, Deciphering post-translational modification codes: FEBS Lett, 2013; 587; 1247-57
26. Woodsmith J, Stelzl U, Studying post-translational modifications with protein interaction networks: Curr Opin Struct Biol, 2014; 24; 34-44
27. Choudhary C, Weinert BT, Nishida Y, The growing landscape of lysine acetylation links metabolism and cell signaling: Nat Rev Mol Cell Biol, 2014; 15; 536-50
Figures
Figure 1. Effect of ANBP on wound healing. (A) Skin wound healing of mice in the ANBP treatment group and the control group at 6 h, 3 d, 7 d and 14 d after wounding. (B) Wound healing rate in the ANBP treatment group and the control group. The wound healing rate of the ANBP treatment group and the control group. Percentage wound healing = (initial wound size – current wound size) ÷ initial wound size ×100%.Figure 2. Workflow for the quantitative acetylome and proteome based on TMT. Three replicates of the global proteome and acetylated proteome were performed in the ANBP treatment group and the control group.Figure 3. Lysine acetylation preferences for significantly changed acetylation sites. Red represents a significant concentration of this amino acid close to the modification site, while green represents a significant reduction in secondary amino acids close to the modification site.Figure 4. Distribution of subcellular location for upregulated and downregulated proteins. (A) Upregulated protein subcellular location in Z6h/C6h; (B) downregulated protein subcellular location in Z6h/C6h.Figure 5. Classification of differentially acetylated sites and heatmap of clustering analysis based on GO enrichment. (A) Classification of differentially acetylated sites Q1, Q2, Q3 and Q4. (B) Heatmap of biological processes for the 4 groups of differentially acetylated sites. (C) Heatmap of cellular components for the 4 groups of differentially acetylated sites. (D) Heatmap of molecular functions for the 4 groups of differentially acetylated sites.Figure 6. Heatmap of clustering analysis based on KEGG pathways and protein domains. (A) Heatmap of KEGG pathways for the 4 groups of differentially acetylated sites. (B) Heatmap of protein domains for the 4 groups of differentially acetylated sites.Figure 7. Protein–protein interaction network of acetylated proteins in periposine-treated AS cells. Tables
Table 1. Comprehensive profiling of the global and acetylated proteomes.Table 2. Number of differentially quantified proteins by comparing ANBP-treated mice with the control.Table 3. The feature sequence around acetylated sites by motif analysis.Table 4. GO annotation for upregulated and downregulated acetylated proteins.Table 1. Comprehensive profiling of the global and acetylated proteomes.Table 2. Number of differentially quantified proteins by comparing ANBP-treated mice with the control.Table 3. The feature sequence around acetylated sites by motif analysis.Table 4. GO annotation for upregulated and downregulated acetylated proteins. In Press
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
08 Mar 2024 : Laboratory Research
Evaluation of Retentive Strength of 50 Endodontically-Treated Single-Rooted Mandibular Second Premolars Res...Med Sci Monit In Press; DOI: 10.12659/MSM.944110
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952