07 July 2020: Lab/In Vitro Research
Induction and Regulation of the Immunoproteasome Subunit β5i (PSMB8) in Laryngeal and Hypopharyngeal Carcinoma Cells
Nan-Xiang Chen1BCEF, Kun Liu1BC, Xuan Liu2A, Xin-Xin Zhang1AD, Dong-yi Han1ADF*DOI: 10.12659/MSM.923621
Med Sci Monit 2020; 26:e923621






Abstract
BACKGROUND: The ubiquitin–proteasome pathway (UPP) is closely associated with the occurrence and progression of cancer, and the 5i immunoproteasome subunit is an important antitumor target in UPP. This study aimed to characterize the regulation of the immunoproteasome subunit β5i (PSMB8) in JHU-011 laryngeal carcinoma cells and FaDu hypopharyngeal carcinoma cells to explore a new target for the treatment of laryngeal and hypopharyngeal carcinomas.
MATERIAL AND METHODS: JHU-011 and FaDu cells were used as effector cells in this study. By means of ⁶⁰Co γ-irradiation, the construction of stable cell lines of the silenced proto-oncogene c-Abl, and the addition of exogenous tyrosine kinase inhibitor (TKI) and activator, the transcription and protein expression levels of PSMB8 and its alternatively spliced isoforms in both cell lines were detected by real-time fluorescence quantitative polymerase chain reaction (RT-PCR) and Western blot.
RESULTS: Ionizing radiation upregulated the transcription level of the alternatively spliced isoform of PSMB8, E2, in both cell lines, thereby upregulating the mRNA and protein levels of PSMB8. The silencing of the proto-oncogene c-Abl and the activation and inhibition of its kinetic kinase product can affect the transcription and protein levels of PSMB8.
CONCLUSIONS: Ionizing radiation can significantly upregulate the mRNA and protein levels of PSMB8, which happens through the upregulation of its splicing isoform E2. The proto-oncogene c-Abl and its kinetic kinase protein product can regulate the transcription and protein expression levels of PSMB8 and its alternatively spliced isoforms.
Keywords: Hypopharyngeal Neoplasms, Laryngeal Neoplasms, Proteasome Inhibitors, Ubiquitin-Specific Proteases, Carcinoma, Immunoproteins, Proteasome Endopeptidase Complex, Proto-Oncogene Mas
Background
Head and neck squamous-cell carcinoma (HNSCC) is a common malignancy in humans, especially in men aged 50–70 years. Its morbidity and mortality rank 6th and 8th among all tumors, respectively [1]. The common HNSCCs are nasopharyngeal carcinoma, oropharyngeal carcinoma, hypopharyngeal carcinoma, laryngeal carcinoma, and nasal cavity, and paranasal sinus carcinoma. Laryngeal carcinoma and hypopharyngeal carcinoma have a poorer prognosis than other HNSCCs. Although the anatomical locations of these 2 are similar, their treatment approaches are not the same due to their different pathogenesis and biological behaviors. With the continuous development of treatments, including surgery, chemoradiotherapy, targeted therapy, and immunotherapy, the quality of life of these patients has significantly improved, but the overall survival has not. Research on new therapeutic targets for laryngeal carcinoma and hypopharyngeal carcinoma is ongoing, and the proteasome is one of the research hotspots.
The ubiquitin–proteasome pathway (UPP) is one of the pathways of selective protein degradation in eukaryotic cells [2]. Protein degradation in UPP entails first linking target proteins to several ubiquitin molecules (i.e., protein ubiquitination), then transporting the ubiquitinated protein to the proteasome for degradation. Proteasomes are divided into constitutive proteasomes and immunoproteasomes. The immunoproteasome is mainly responsible for antigen presentation, T cell differentiation, regulation of cytokines, antioxidative stress, and other functions. It is not only involved in important metabolic processes in the human body but also closely involved in the occurrence and progression of tumors [3].
Although a comprehensive treatment has been adopted for common HNSCCs with a poor prognosis, such as hypopharyngeal carcinoma and laryngeal carcinoma, the survival of patients is still very limited. Exploring new treatment methods or therapeutic targets is the research direction for breaking the bottleneck of therapeutic effects. Using proteasomes as a potential therapeutic target is based on research by King et al. [4], which indicated that inhibitors of cyclin-dependent kinases could cause cell cycle arrest and cell apoptosis if these inhibitors were not promptly degraded and inactivated via specific degradation pathways mediated by UPP. Therefore, it has been speculated that proteasome inhibitors (PIs) may be able to inhibit the proliferation of tumor cells. Further studies have found that the expression and activity of proteasomes are much higher in tumor tissue than in normal tissue. The difference is due to a higher need for proteasomes in tumor cells, wherein proteasomes are involved in the removal of proteins with translational errors or spatial conformational abnormalities, caused by genetic instability of tumor cells and their disordered and rapid proliferation [5]. Therefore, compared with normal cells, tumor cells are more sensitive to PIs. PIs act on proteasomes in the UPP of tumor cells, affecting intracellular signaling pathways and playing a specific role in inhibiting the proliferation of tumor cells or inducing tumor cell apoptosis.
The structure of the proteasome in eukaryotic cells is shown in Figure 1. The 2 outer rings are each composed of 7 different α subunits, and the 2 inner rings are composed of 7 different β subunits, among which activities of 3 β subunits have been identified. The β1 subunit has a caspase-like activity; the β2 subunit has a trypsin-like activity; and the β5 subunit has a chymotrypsin-like activity [6]. In higher eukaryotes, these 3 subunits can be induced by cytokines (such as IFN-γ) and converted into 3 active forms, namely, β1i (LMP2, PSMB9), β2i (MECL-1, PSMB10), and β5i (LMP7, PSMB8), which replace β1, β2, and β5, respectively, to form immunoproteasomes [7]. The chymotrypsin region is the main functional region involved in cell growth and proliferation, and PSMB8 mRNA is alternatively spliced, giving rise to 2 protein isoforms, E1 and E2. Therefore, the activity of PSMB8 is more easily regulated by the ratio of the isoforms, and PSMB8 is the site that can easily affect the proteasome activity. At present, 3 PIs, i.e., bortezomib, carfilzomib, and ixazomib, are mainly used in the clinic to treat multiple myeloma by targeting and inhibiting PSMB8 (Table 1). If the regulation of PSMB8 in laryngeal and hypopharyngeal carcinomas is clarified, the data could provide evidence and clues for the exploration of PSMB8 as an important new target for the treatment of these HNSCCs.
In addition, studies have shown that ionizing radiation affects the 19S and 20S proteasome subunits in model organisms [8]. Since radiotherapy is widely used in the treatment of HNSCCs, the induction effect of ionizing radiation on laryngeal and hypopharyngeal carcinomas is worth studying. Ionizing radiation can not only produce a large amount of oxygen free radicals in cells and induce cell apoptosis and DNA damage repair [9] but also enhance the cytotoxic effects of targeted drugs on tumor cells [10]. A recent study has indicated that the non-receptor tyrosine kinase encoded by the proto-oncogene c-Abl is associated with radiotherapy resistance, and imatinib, which targets this tyrosine kinase, could radiosensitize HNSCC cells [11]. Meanwhile, the c-Abl-encoded non-receptor tyrosine kinase regulates UPP by controlling the degradation of proteasome subunits [12,13]. Therefore, research on the regulation of the c-Abl-encoded tyrosine kinase in proteasomes may lead to the discovery of new targeted drugs, which can be combined with radiotherapy to compensate for the shortage of targeted drugs for HNSCC and for the development of drug resistance in patients after initial susceptibility to targeted drugs.
However, in the studies on proteasomes and head and neck tumors, the regulatory processes of proteasomes in laryngeal carcinoma and hypopharyngeal carcinoma cells have not been reported. Therefore, we based this study on the dominant role of the β5i (LMP7, PSMB8) subunit in immunoproteasome function; on the data obtained by using PSMB8-targeting antitumor PIs; on the important role of radiotherapy in the treatment of laryngeal and hypopharyngeal carcinomas; and on the possible synergistic effect of combination of PIs with radiotherapy. This study modulated the c-Abl gene expression and the activity of its encoded tyrosine kinase through both endogenous (silencing of c-Abl gene) and exogenous means (ionizing radiation, inhibition and activation of tyrosine kinase), and observed the regulation of the mRNA and protein levels of PSMB8 in JHU-011 laryngeal carcinoma cells and FaDu hypopharyngeal carcinoma cells, in order to explore the potential of the proteasome as a therapeutic target for laryngeal carcinoma and hypopharyngeal carcinoma.
Material and Methods
Material
CELL LINES AND CULTURE CONDITIONS:
JHU-011 and FaDu cells (a gift from Dr. Califano’s laboratory, Johns Hopkins University) were cultured in RPM-1640 medium (GIBCO). MCF-7 human breast carcinoma cells and human embryonic kidney 293T (HEK293T) cells (Beijing Institute of Biotechnology, Beijing 100850, China) were cultured in Dulbecco’s modified Eagle’s medium (DMEM, GIBCO). Heat-inactivated fetal bovine serum (HyClone) was added into all media. Culture conditions: 37°C, 8% CO2, pH 7.2–7.4, and aseptic culture at constant temperature.
INDUCTION EXPERIMENT BY IONIZING γ-RADIATION:
JHU-011 and FaDu cells were divided into 5 groups according to the dose (0 Gy, 2.5 Gy, 5 Gy, 10 Gy, and 20 Gy). The culture dish was irradiated with γ-rays at room temperature, and the cells were continuously cultured for 12 h in the incubator after radiation. RNA was then extracted from the cells of each group and reverse-transcribed into cDNA. RT-PCR was performed to find the dose that achieved the maximum transcriptional induction of PSMB8. Then, the JHU-011 cells and FaDu cells were irradiated at this dose. After radiation, RT-PCR and Western blot were used to detect the changes in the transcription and protein expression levels of PSMB8 (and its 2 alternatively spliced isoforms E1 and E2) in FaDu and JHU-011 cells. A fixed 60Co γ-ray source was used, and the total radiation dose was adjusted by changing the duration of radiation.
DETECTION OF THE RRANSCRIPTION LEVELS OF PSMB8 AND ITS ALTERNATIVELY SPLICED ISOFORMS USING RT-PCR:
The RNA of the cells of each group was extracted. The RNA was reverse-transcribed into cDNA and added into the reaction system (cDNA 1 μl, 2× iQTM SYBR Green Mix 10 μl, forward primer 1 μl, reverse primer 1 μl, ddH2O 7 μl) for RT-PCR. β-Actin was used as the internal reference. The primers were as follows:
PSMB8 primers:
β-Actin primers:
The primers for alternatively spliced isoforms E1 and E2:
The RT-PCR was performed using the Bio-Rad iQ5 Real-Time PCR System. The samples were denatured at 95°C for 10 min, followed by 45 cycles of amplification (95°C, 10 s for denaturation; 60°C, 30 s for annealing). Each sample was run in triplicate, and the data were analyzed using the ΔΔCt method.
PROTEIN EXPRESSION OF PSMB8 DETECTED BY WESTERN BLOT:
The cells of each group were lysed, 200 μl of single cell-lysate was added to each culture dish, placed in ice-water for 30 min, and then centrifuged at 12, 000 rpm/min at 4°C for 10 min. After lysis, the Bio-Rad spectrophotometer was used to measure the absorbance of the cells at 595 nm. Then the protein concentration in the lysate of each group was balanced according to the absorbance. Samples were boiled at 100°C for 5 min, and 10 μl of each sample was used for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (polyacrylamide gel concentration: 12%; β-actin as internal control). The polyacrylamide gel was first placed on the poly(vinylidene fluoride) (PVDF) membrane. Then, the stack of filter paper-gel-PVDF membrane–filter paper was placed on a semidry transfer unit for 2 h at 18 V. After blocking in skim milk, the sample was incubated with the properly diluted primary antibody at room temperature for 1 h. After washing, the sample was incubated with properly diluted secondary antibody at room temperature for 1 h. The primary antibody was anti-goat LMP7, the secondary antibody was rabbit anti-goat, and the internal reference was HRP-β-actin. The membranes were exposed to X-ray film (Fuji) and developed with the electrochemiluminescence (ECL) system (Perkin Elmer). After scanning the X-ray films, ImageJ software was used for grayscale analysis, and the relative protein expression levels were calculated based on the grayscale values.
CONSTRUCTION OF C-ABL-SHRNA/C-ABL-ARG PLASMID:
According to the nucleotide sequences of c-Abl and Arg in GenBank (National Center for Biotechnology Information), starting from 25 nucleotides downstream of the initiation codon ATG, we searched for 21 nucleotides that conformed to AA (N19) and designed the forward oligo and reverse oligo. c-Abl-short hairpin RNA (shRNA) had a silencing effect on the target gene c-Abl, and c-Abl-Arg did not have a silencing effect.
The sequences of the forward oligo and reverse oligo of c-Abl were as follows:
Arg was used as the negative control of c-Abl. The sequences of the forward oligo and reverse oligo were as follows:
The pLKO.1-TCR vector was digested with AgeI (NEB, R0552S), and the enzyme digestion system was separated by electrophoresis on a 0.8% agarose gel. The recovered products were purified and then digested with EcoRI (NEB, R0101S). After separating the enzyme digestion system, the vector fragment (approximately 7 kb) was recovered. The double-stranded oligonucleotides of c-Abl and Arg were cloned into the recovered pLKO.1 vector fragment after the double enzyme digestion with AgeI and EcoRI, then transformed into DH5α competent cells. Five to 10 monoclonal colonies were screened in Luria-Bertani medium with ampicillin selection and incubated at 37°C for 12 h on a shaker at 200 rpm/min, followed by extracting the resistance-conferring plasmids. The primer 5′-CAAGGCTGTTAGAGAGATAATTGGA-3′ was used for sequencing.
COLLECTING VIRUSES INFECTED CELLS AND SCREENING THE STABLE CELL LINES:
One microgram of the constructed pLKO.1-c-Abl-shRNA plasmid, 750 ng of psPAX2 plasmid, and 250 ng of pMD2. G plasmid were added into an Eppendorf tube, and the serum-free OPTI-MEM medium was added up to a volume of 20 μl. After it was mixed with the transfection reagent, the solution was left for 30 min and then added to 293T cells for infection. After 48 h, the culture medium was changed, and the supernatant was collected. FaDu cells and JHU-011 cells were introduced into 6-well plates and cultured at approximately 2×105 cells/well, with the addition of puromycin (0.5 μg/ml). The above collected supernatant was added with polybrene (final concentration 8 μg/ml) into 6-well plates to infect the cells for 24 h. By often refreshing the culture medium containing puromycin, the cell lines stably expressing pLKO.1-c-Abl-shRNA were screened.
The cell lines of hypopharyngeal carcinoma and laryngeal carcinoma with stable expression of pLKO.1-c-Abl-Arg were prepared and screened using the same method.
ENDOGENOUS SILENCING OF THE TARGET GENE C-ABL:
JHU-011 cells and FaDu cells were divided into 3 groups: the normal control group (Nor) without lentivirus infection, the pLKO.1-c-Abl-Arg–transfected group (c-Abl-Arg), and the PLKO.1-c-Abl-shRNA-transfected group (c-Abl-shRNA). Each group was subdivided into 2 subgroups, one with and the other without IFN-γ induction. The subgroups with IFN-γ were stimulated for 24 h at a final concentration of 150 U/μl, followed by RT-PCR and Western blot.
GROUPING FOR THE EXOGENOUS ACTIVATION/INHIBITION OF TYROSINE KINASE:
JHU-011 cells and FaDu cells were inoculated in 10-cm culture dishes and divided into 5 groups according to the medium: normal 1640 medium without any inhibitor/activator (IFN-γ−/Nilo−), medium with nilotinib but without exogenous H-IFN-γ (IFN-γ−/Nilo+), medium with exogenous H-IFN-γ but without nilotinib (IFN-γ+/Nilo−), medium with both nilotinib and exogenous H-IFN-γ (IFN-γ+/Nilo+), and medium with only exogenous DPH (DPH+). In addition, the cells in the medium with both nilotinib and exogenous IFN-γ were first pretreated with nilotinib for 24 h, followed by the addition of IFN-γ, then were cultured for 24 h before RT-PCR and Western blot. The cells in DPH group were cultured for 48 h before tests. The final concentrations of nilotinib, IFN-γ, and DPH in the culture medium were 5 mM, 150 μ/ml, and 10 mM, respectively.
Results
Material
EFFECT OF IONIZING γ-RADIATION ON THE MRNA AND PROTEIN LEVELS OF PSMB8 AND ITS 2 ALTERNATIVELY SPLICED ISOFORMS:
The involvement of proteasomes in the oxidative stress response induced by ionizing radiation has become increasingly evident in recent studies [14,15]. Radiotherapy is an important treatment modality for patients with laryngeal carcinoma and hypopharyngeal carcinoma. To study the role of acute ionizing radiation in UPP and the regulatory effect of acute ionizing radiation on PSMB8, we first determined the dose of radiation that maximized the transcriptional induction of PSMB8. The results showed that 60Co γ-irradiation upregulated the transcription level of PSMB8 in FaDu cells and JHU-011 cells, and the maximum upregulation was achieved at a dose of 10 Gy (Figure 2A). In addition, we measured the Western blot gray value of PSMB8 using the ImageJ software. The histogram (Figure 2B) shows that following 10 Gy irradiation, the protein expression of PSMB8 was significantly upregulated in FaDu cells (** P<0.01) and JHU-011 cells (*** P<0.001). 60Co ionizing γ-radiation significantly upregulated the transcription level of the alternatively spliced isoform E2 of PSMB8 (** P<0.01) (Figure 2C).
CONSTRUCTION OF THE PLKO.1-C-ABL-SHRNA AND PLKO.1-C-ABL-ARG PLASMIDS: The lentiviral vector constructed in this study was composed of 3 different plasmids, pLKO.1-TRC, psPAX2, and pMD2. G. The pLKO.1-TRC vector was used to express the RNA interference (RNAi) fragment of c-Abl (Figure 2D), and the psPAX2 vector and pMD2. G vector were complementary vectors.
EFFECT OF TARGETED SILENCING OF THE C-ABL GENE ON PSMB8 AND ITS ALTERNATIVE SPLICING ISOFORMS: This study showed that IFN-γ significantly induced the transcription of PSMB8 (** P<0.01). Compared with that in the pLKO.1-c-Abl-Arg group, targeted silencing of the c-Abl gene by stably expressing pLKO.1-c-Abl-shRNA in cells downregulated the IFN-γ-induced transcription level of PSMB8 in the laryngeal and hypopharyngeal carcinoma cells (Figure 3A), thus downregulating the PSMB8 protein (** P<0.01) (Figure 3B, 3C), and the changes in the mRNA and protein expression were consistent. In addition, RT-PCR showed that the targeted silencing of the c-Abl gene changed the ratio between 2 alternatively spliced isoforms of PSMB8 in FaDu and JHU-011 cells and downregulate the mRNA level of E2 (Figure 3D, 3E).
EFFECT OF EXOGENOUS TYROSINE KINASE INHIBITOR OR ACTIVATOR ON THE MRNA AND PROTEIN LEVELS OF PSMB8 AND ITS ALTERNATIVELY SPLICED ISOFORMS:
We chose nilotinib to exogenously inhibit the tyrosine kinase encoded by the proto-oncogene c-Abl, and exogenous activation was performed using the c-Abl activator DPH. Nilotinib is a highly selective inhibitor of BCR-ABL kinase. We observed that nilotinib, an exogenous TKI, downregulated the transcription level of PSMB8 in the IFN-γ-induced JHU-011 cells and FaDu cells (Figure 4A). In addition, PSMB8 protein was downregulated (Figure 4B. 4C). The ratio between alternatively spliced isoforms was also detected. It was found that the exogenous inhibitory effect of nilotinib mainly caused the relative percentage of E2 to decrease (Figure 4D. 4E). Compared with the control group, DPH upregulated the mRNA and protein levels of PSMB8 in JHU-011 cells and FaDu cells (Figure 4F).
Discussion
UPP is closely involved in the cell cycle, cell differentiation, cell proliferation, injury repair, antigen presentation, and various intracellular signal pathways. Currently, the mechanisms by which UPP participates in tumor development are as follows: the activity of p53 protein is regulated by murine double minute 2 (MDM2); the function of MDM2 is similar to that of ligase E3 in UPP, which can promote the degradation of p53 protein through UPP, thus affecting the antitumor effect of p53 and promoting the occurrence of tumor [16,17]. The nuclear transcription factor NF-κB is stably expressed in a variety of tumors, especially in the head and neck tumors [18,19]. Most NF-κB dimers bind to the inhibitor IκB and remain in an inactive state. When cells are stimulated by certain external factors, the inhibitory factor IκB is degraded by UPP, thereby increasing the activity of NF-κB. UPP can also promote the degradation of inhibitor IκB precursors (P150, P100) and activate the NF-κB signaling pathway [20]. Therefore, the important intracellular signal transduction pathways associated with tumor development are regulated by the UPP.
In this study, ionizing γ-radiation was used since this radiation has an effect on proteasome activity [21,22]. Acute ionizing radiation can cause a large amount of oxygen free radical accumulation, cell apoptosis, repair of DNA damage, and changes in the expression levels of many genes [9]. Second, radiotherapy plays an important role in the treatment of head and neck tumors. Radiotherapy not only can directly kill local carcinoma cells but also may result in the abscopal effect, which could increase the tumor cell sensitivity to immunotherapies [23]. PSMB8 is the main target of commercially available PIs. The results of this study indicated that ionizing radiation had an upregulating effect on PSMB8 in laryngeal and hypopharyngeal carcinoma cells, which was mainly due to the regulation of the alternatively spliced isoform E2. The induction and enhancement effects of ionizing radiation on PSMB8 can increase the level of recognition of PSMB8 by potential PIs, thus resulting in a synergistic effect of combined radiotherapy and PIs. More importantly, since immunoproteasomes are involved in antigen presentation in tumor cells, the regulatory effect of ionizing radiation on proteasomes can enhance the immunogenicity or antigen-presenting function of tumor cells, which may thus be more easily recognized and killed by T cells. The results of this study suggest that the effect of γ-irradiation on PSMB8 might be realized by changing the ratio between PSMB8 alternatively spliced isoforms, and the γ-irradiation could regulate the transcription and protein expression levels of PSMB8 and affect its activity. We speculate that the possible mechanisms are as follows: 1. Ionizing radiation affects the upstream pathway of proteasome expression, such as the IFN-γ signal transduction pathway, which is sensitive to radiation, thereby altering the transcription and expression levels of proteasome members. 2. Radiation may affect different promoters in the transcription of PSMB8, resulting in inconsistent expression levels of the initial exons and the differential expression between the 2 alternatively spliced isoforms. 3. Radiation activates certain phosphorylation pathways, thereby promoting the transcription and assembly processes of PSMB8.
To further study the regulation of PSMB8, we used the proto-oncogene c-Abl as the key. C-Abl is a homologous gene of the proto-oncogene v-Abl [24]. It encodes a non-receptor tyrosine kinase that can catalyze the phosphorylation of tyrosine residues in many substrate proteins. It has been confirmed that the c-Abl gene can regulate proteasome subunits, and the c-Abl-encoded tyrosine kinase may be involved in radiation resistance of cancer cells; however, the regulatory effects of c-Abl on proteasome subunits have not been reported in laryngeal and hypopharyngeal cancer cells. In this study, c-Abl was used as the target gene to construct the pLKO.1-c-Abl-shRNA lentiviral vector, and endogenous silencing of c-Abl was done by the double-stranded RNA–mediated RNAi process. The results suggest that endogenous silencing of the c-Abl gene can downregulate the transcription and protein expression levels of PSMB8 mainly by affecting the transcription of isoform E2. We speculate that isoform E2 plays a major role in the activity of the immunoproteasome. The constructed pLKO.1-c-Abl-shRNA plasmid and the stably transfected cell lines allowed us to knock down the target gene c-Abl, which lays a foundation for further study on the regulatory mechanisms of proteasomes and other biological effects in laryngeal carcinoma and hypopharyngeal carcinoma cells.
The transformation process of normal cells to tumor cells in the head and neck is closely related to the activation of tyrosine kinases. For example, epidermal growth factor receptor (EGFR) belongs to the tyrosine kinase receptor family. To further clarify the roles of the upstream and downstream proteins of PSMB8, we further activated and inhibited the expression product of c-Abl. Unlike EGFR, the protein encoded by c-Abl is a non-receptor tyrosine kinase. Small-molecule TKIs, such as imatinib, dasatinib, and nilotinib, can exert their inhibitory effects by inhibiting BCR-Abl fusion protein. We chose the TKI nilotinib and activator DPH and observed whether their exogenous effect on c-Abl gene expression products would affect PSMB8. On the one hand, the results confirmed consistent regulatory effects of the c-Abl gene and its protein product on the upstream and downstream effectors of PSMB8 in laryngeal and hypopharyngeal carcinoma cells, showing that the induction of c-Abl in proteasomes was achieved through its encoded tyrosine kinase rather than through other pathways. On the other hand, the upregulating effect of c-Abl and its tyrosine kinase on isoform E2 of PSMB8 further confirmed the importance of E2 in the regulation of PSMB8, suggesting that the c-Abl pathway can be used as a potential inhibition pathway and E2 can be used as a potential inhibition target.
Bortezomib, a specific PI that has been used in clinical practice, was approved by the US Food and Drug Administration in 2003 for the treatment of multiple myeloma. The drug binds to the 20S proteasome subunit β5i and 26S proteasome subunit β1i and achieves its antitumor effects by inhibiting the decomposition of specific proteins related to tumor suppressor genes in UPP [25,26]. For head and neck tumors, bortezomib does not significantly inhibit proliferation-related pathways, such as STAT3, and has no significant effect on the expression of EGFR; therefore, it has poor efficacy in HNSCC in drug experiments and clinical applications [27,28]. Nevertheless, the role of PIs in the treatment of other tumors still provides new thinking for the treatment of head and neck tumors, and some studies are currently underway [29–31]. This study, for the first time, elucidated the effect of proteasome inhibition on the UPP and PSMB8 in laryngeal and hypopharyngeal carcinoma cells. The upregulation of PSMB8 by ionizing radiation (radiotherapy) may synergistically enhance the targeting of PSMB8 by PIs, and the regulatory effects of the c-Abl gene and its expression product on isoform E2 confirmed the importance of E2 in the regulation of PSMB8. These results can provide some references for exploring new therapeutic targets for HNSCCs and facilitate the application of PIs in the treatment of HNSCCs.
Conclusions
Ionizing radiation can upregulate the mRNA and protein levels of PSMB8 in FaDu cells and JHU-011 cells, which happens through the upregulation of its splicing isoform E2. The proto-oncogene c-Abl and its kinase protein product can regulate the transcription and protein expression levels of PSMB8. The findings of this study imply that PSMB8 may be a potential research target in the treatment of laryngeal and hypopharyngeal carcinoma.
Figures
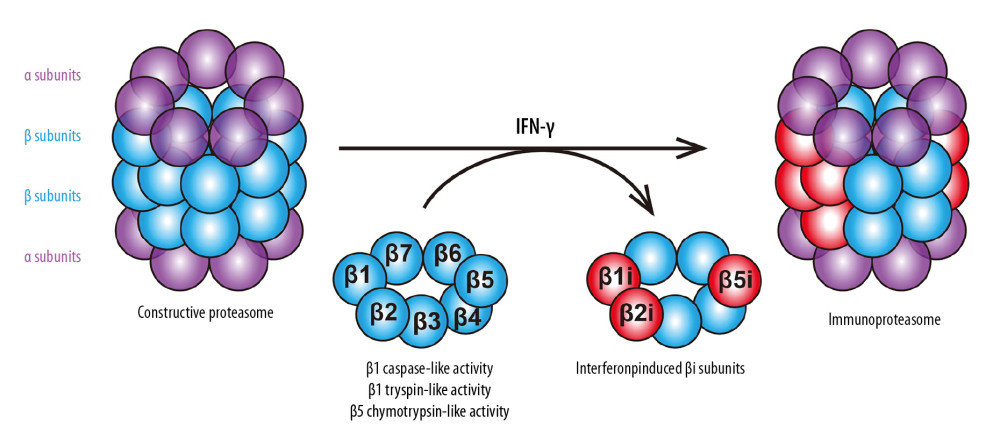 Figure 1. Upon stimulation with cytokines (e.g., IFN-γ), the constitutively expressed β-subunits (β1, β2, and β5) of the 20S constructive proteasome are replaced by their inducible counterparts, β1i, β2i, and β5i, respectively, which form the 20S immunoproteasome (i20S).
Figure 1. Upon stimulation with cytokines (e.g., IFN-γ), the constitutively expressed β-subunits (β1, β2, and β5) of the 20S constructive proteasome are replaced by their inducible counterparts, β1i, β2i, and β5i, respectively, which form the 20S immunoproteasome (i20S). 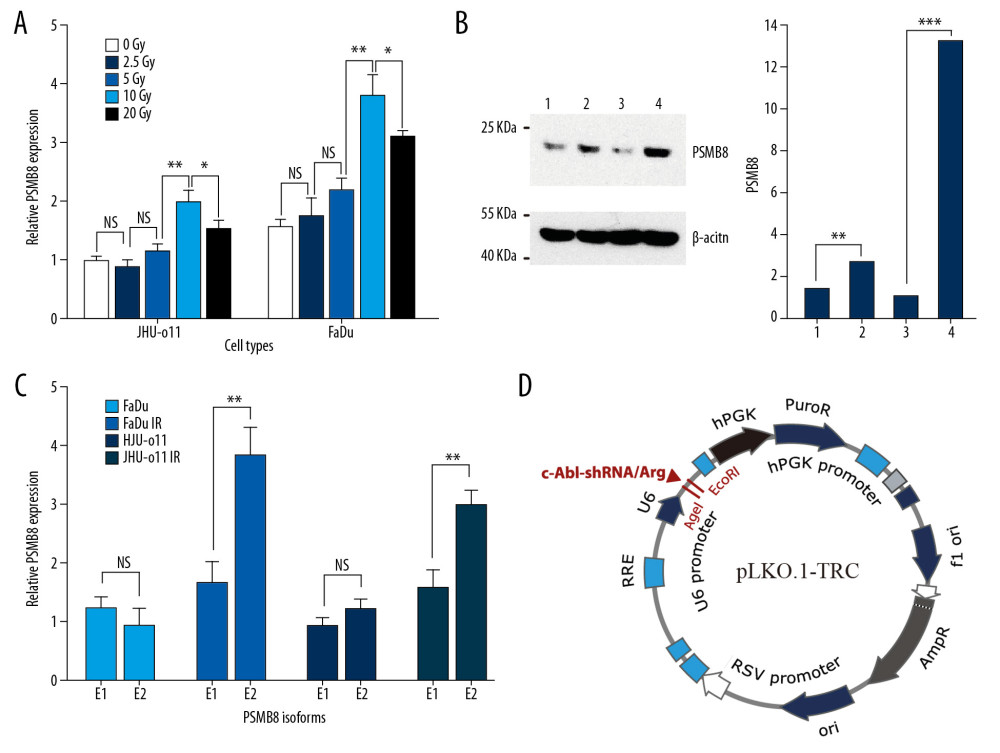 Figure 2. (A) JHU-011 cells, FaDu cells were divided into 5 groups according to the dose gradient (0 Gy, 2.5 Gy, 5 Gy, 10 Gy, 20 Gy), and γ-irradiation was carried out at room temperature. After irradiation, the cells were cultured for 12 h, and then the RT-PCR was performed to detect the transcription level of PSMB8. The dose of 10 Gy showed the most significant induction effect on all 3 types of the cells (** P<0.01). (B) Western blot of PSMB8 in FaDu cells and JHU-011 cells. Lanes: 1, FaDu cells without irradiation; 2, FaDu cells with irradiation; 3, JHU-011 cells without irradiation; 4, JHU-011 cells with irradiation. (C) The dose of 10 Gy was used to irradiate the JHU-011 cells and FaDu cells, and the relative ratio between isoforms E1 and E2 was detected by RT-PCR. The percentage of E2 was significantly upregulated by irradiation in FaDu cells (** P<0.01) and JHU-011 cells (** P<0.01). Each group included 3 independent repeats. Quantitative data are expressed as the mean±standard deviation (SD). (D) Plasmid pLKO.1-TRC contained the insertion site of the shRNA, located between the U6 promoter and cPPT, which originally was a stuffer of approximately 1.9 kb long. After digestion with AgeI and EcoRI, the designed double-stranded shRNA oligos (c-Abl-shRNA and c-Abl-Arg) were inserted into the gap between the AgeI and EcoRI sites to replace the stuffer, yielding the vector.
Figure 2. (A) JHU-011 cells, FaDu cells were divided into 5 groups according to the dose gradient (0 Gy, 2.5 Gy, 5 Gy, 10 Gy, 20 Gy), and γ-irradiation was carried out at room temperature. After irradiation, the cells were cultured for 12 h, and then the RT-PCR was performed to detect the transcription level of PSMB8. The dose of 10 Gy showed the most significant induction effect on all 3 types of the cells (** P<0.01). (B) Western blot of PSMB8 in FaDu cells and JHU-011 cells. Lanes: 1, FaDu cells without irradiation; 2, FaDu cells with irradiation; 3, JHU-011 cells without irradiation; 4, JHU-011 cells with irradiation. (C) The dose of 10 Gy was used to irradiate the JHU-011 cells and FaDu cells, and the relative ratio between isoforms E1 and E2 was detected by RT-PCR. The percentage of E2 was significantly upregulated by irradiation in FaDu cells (** P<0.01) and JHU-011 cells (** P<0.01). Each group included 3 independent repeats. Quantitative data are expressed as the mean±standard deviation (SD). (D) Plasmid pLKO.1-TRC contained the insertion site of the shRNA, located between the U6 promoter and cPPT, which originally was a stuffer of approximately 1.9 kb long. After digestion with AgeI and EcoRI, the designed double-stranded shRNA oligos (c-Abl-shRNA and c-Abl-Arg) were inserted into the gap between the AgeI and EcoRI sites to replace the stuffer, yielding the vector. 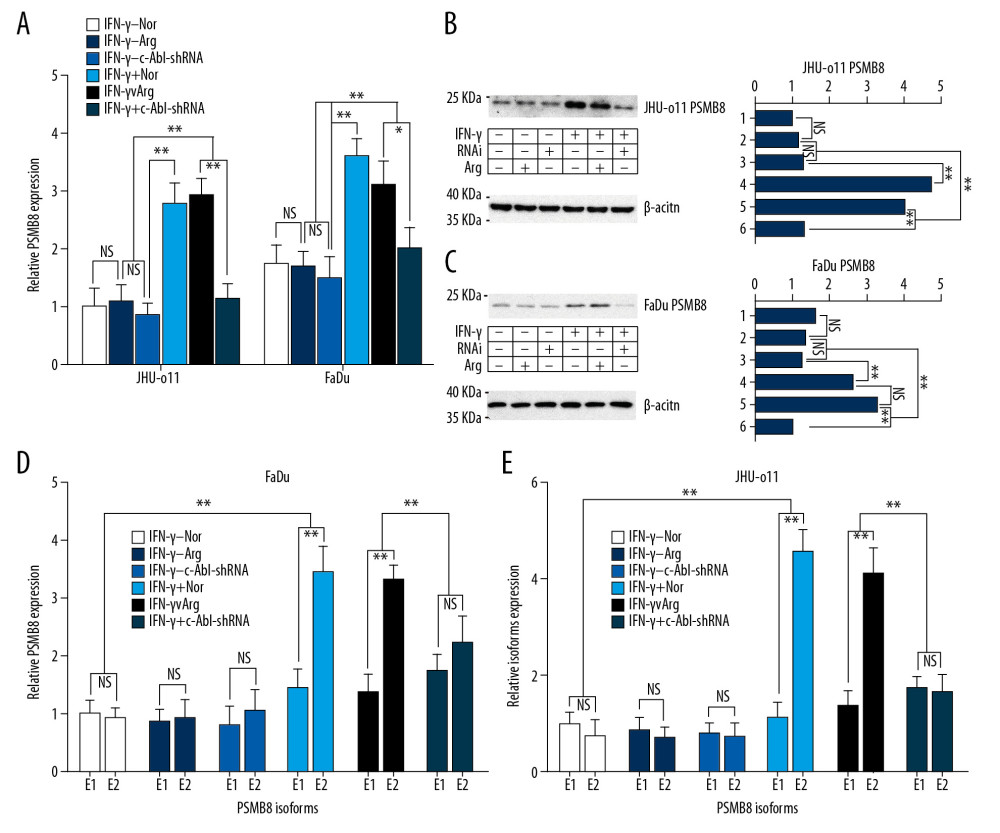 Figure 3. Effect of targeted silencing of the c-Abl gene on the mRNA and protein levels of PSMB8 and its alternatively spliced isoforms. (A) JHU-011 cells and FaDu cells were divided into 6 groups (described previously). In the 3 groups with IFN-γ (final concentration of 150 U/μl), the transcription of PSMB8 in the IFN-γ+/Nor group and the IFN-γ+/Arg group was significantly upregulated but was inhibited in the IFN-γ+/c-Abl-shRNA group due to the targeted silencing of the c-Abl gene (** P<0.01). (B) PSMB8 protein expression levels were detected in JHU-011 cells by Western blotting, and the ImageJ software was used to determine the gray value and drew a histogram. Lanes 1–6 represent IFN-γ−/RNAi−/Arg−, IFN-γ−/RNAi−/Arg+, IFN-γ−/RNAi+/Arg−, IFN-γ+/RNAi−/Arg−, IFN-γ+/RNAi−/Arg+, and IFN-γ+/RNAi+/Arg−, respectively. The results show that the PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg− group (** P<0.01). (C) The PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg- group of FaDu cells (** P <0.01). (D) FaDu cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group. The transcription level of isoform E2 was significantly upregulated by IFN-γ (** P<0.01). The targeted silencing of c-Abl gene (IFN-γ+/c-Abl-shRNA) downregulated the transcription of E2 (** P<0.01). (E) JHU-011 cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group, and the results were consistent with those of FaDu cells (** P<0.01). Each group contained 3 independent replicates. Quantitative data are expressed as the mean±SD.
Figure 3. Effect of targeted silencing of the c-Abl gene on the mRNA and protein levels of PSMB8 and its alternatively spliced isoforms. (A) JHU-011 cells and FaDu cells were divided into 6 groups (described previously). In the 3 groups with IFN-γ (final concentration of 150 U/μl), the transcription of PSMB8 in the IFN-γ+/Nor group and the IFN-γ+/Arg group was significantly upregulated but was inhibited in the IFN-γ+/c-Abl-shRNA group due to the targeted silencing of the c-Abl gene (** P<0.01). (B) PSMB8 protein expression levels were detected in JHU-011 cells by Western blotting, and the ImageJ software was used to determine the gray value and drew a histogram. Lanes 1–6 represent IFN-γ−/RNAi−/Arg−, IFN-γ−/RNAi−/Arg+, IFN-γ−/RNAi+/Arg−, IFN-γ+/RNAi−/Arg−, IFN-γ+/RNAi−/Arg+, and IFN-γ+/RNAi+/Arg−, respectively. The results show that the PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg− group (** P<0.01). (C) The PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg- group of FaDu cells (** P <0.01). (D) FaDu cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group. The transcription level of isoform E2 was significantly upregulated by IFN-γ (** P<0.01). The targeted silencing of c-Abl gene (IFN-γ+/c-Abl-shRNA) downregulated the transcription of E2 (** P<0.01). (E) JHU-011 cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group, and the results were consistent with those of FaDu cells (** P<0.01). Each group contained 3 independent replicates. Quantitative data are expressed as the mean±SD. 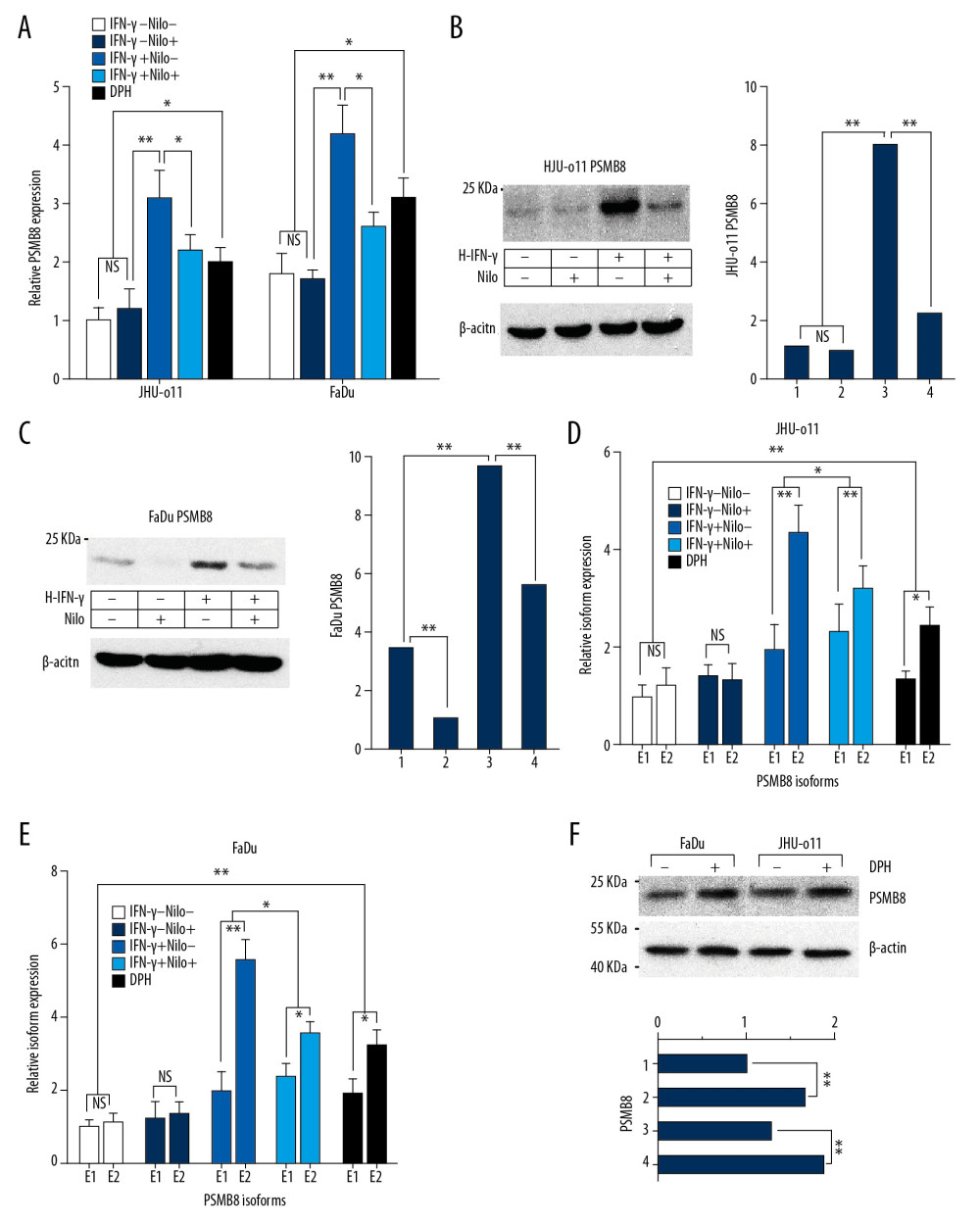 Figure 4. Regulation of tyrosine kinase activity on the transcription and expression levels of PSMB8 and its alternatively spliced isoforms (A) JHU-011 cells and FaDu cells were divided into 5 groups. In the IFN-γ+/Nilo+ group, nilotinib was first added and the cells were incubated for 24 h. Then IFN-γ was added and the cells were cultured for another 24 h. In the IFN-γ+/Nilo− group, the transcription level of PSMB8 was significantly upregulated compared to that in the control group (IFN-γ−/Nilo− group) (** P<0.01), while the transcription of PSMB8 in the IFN-γ−/Nilo+ group was inhibited (* P<0.05). (B) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated compared to that in the IFN-γ+/Nilo− group in FaDu cells (** P<0.01). (C) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated in JHU-011 cells (** P<0.01). (D) Nilotinib downregulated the induction effect of IFN-γ on isoform E2, and the percentage of isoform E2 was reduced (* P<0.05). In the DPH+ group, the transcription level of isoform E2 was higher than that in the control group (IFN-γ−/Nilo− group) (** P<0.01). Quantitative data are expressed as the mean±SD. (E) In FaDu cells, the regulation of nilotinib on the transcription of E2 was consistent with the results in JHU-011 cells (* P<0.05). (F) In the DPH+ group, protein expression of PSMB8 in JHU-011 cells and FaDu cells was elevated compared to that in the blank control group (** P<0.01). Each group contained 3 independent replicates.
Figure 4. Regulation of tyrosine kinase activity on the transcription and expression levels of PSMB8 and its alternatively spliced isoforms (A) JHU-011 cells and FaDu cells were divided into 5 groups. In the IFN-γ+/Nilo+ group, nilotinib was first added and the cells were incubated for 24 h. Then IFN-γ was added and the cells were cultured for another 24 h. In the IFN-γ+/Nilo− group, the transcription level of PSMB8 was significantly upregulated compared to that in the control group (IFN-γ−/Nilo− group) (** P<0.01), while the transcription of PSMB8 in the IFN-γ−/Nilo+ group was inhibited (* P<0.05). (B) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated compared to that in the IFN-γ+/Nilo− group in FaDu cells (** P<0.01). (C) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated in JHU-011 cells (** P<0.01). (D) Nilotinib downregulated the induction effect of IFN-γ on isoform E2, and the percentage of isoform E2 was reduced (* P<0.05). In the DPH+ group, the transcription level of isoform E2 was higher than that in the control group (IFN-γ−/Nilo− group) (** P<0.01). Quantitative data are expressed as the mean±SD. (E) In FaDu cells, the regulation of nilotinib on the transcription of E2 was consistent with the results in JHU-011 cells (* P<0.05). (F) In the DPH+ group, protein expression of PSMB8 in JHU-011 cells and FaDu cells was elevated compared to that in the blank control group (** P<0.01). Each group contained 3 independent replicates. 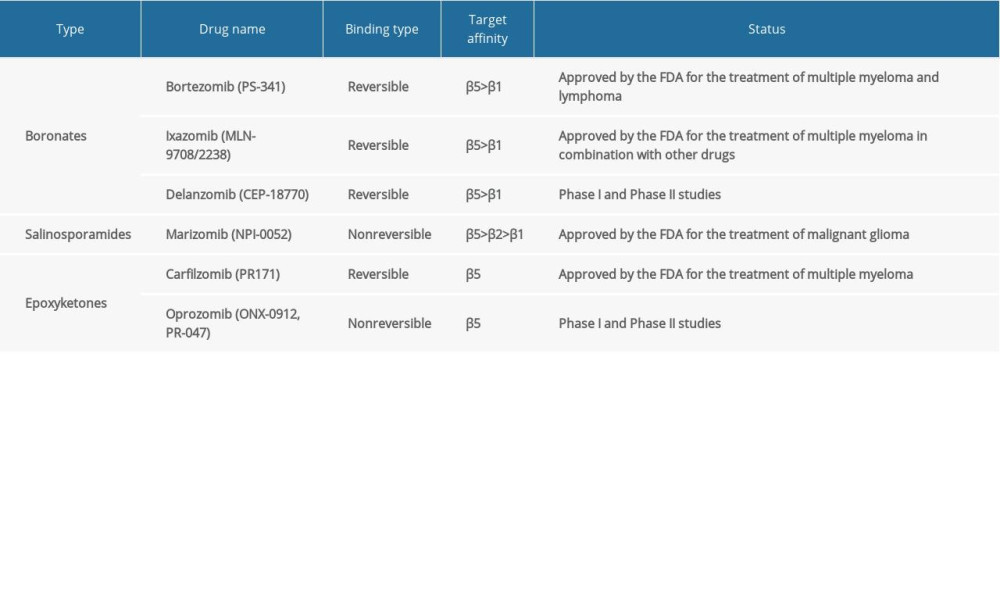
References
1. Parfenov M, Pedamallu CS, Gehlenborg N, Characterization of HPV and host genome interactions in primary head and neck cancers: Proc Natl Acad Sci USA, 2014; 111; 15544-49
2. Mukhopadhyay D, Riezman H, Proteasome-independent functions of ubiquitin in endocytosis and signaling: Science, 2007; 315; 201-5
3. Angeles A, Fung G, Luo H, Immune and non-immune functions of the immunoproteasome: Front Biosci (Landmark Ed), 2012; 17; 1904-16
4. King RW, Deshaies RJ, Peters JM, Kirschner MW, How proteolysis drives the cell cycle: Science, 1996; 274; 1652-59
5. Chen L, Madura K, Increased proteasome activity, ubiquitin-conjugating enzymes, and eEF1A translation factor detected in breast cancer tissue: Cancer Res, 2005; 65; 5599-606
6. Orlowski RZ, Kuhn DJ, Proteasome inhibitors in cancer therapy: Lessons from the first decade: Clin Cancer Res, 2008; 14; 1649-57
7. Ferrington DA, Gregerson DS, Immunoproteasomes: Structure, function, and antigen presentation: Prog Mol Biol Transl Sci, 2012; 109; 75-112
8. Cerqueira PG, Passos-Silva DG, Vieira-da-Rocha JP: Mol Biochem Parasitol, 2017; 212; 55-67
9. Azzam EI, Jay-Gerin JP, Pain D, Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury: Cancer Lett, 2012; 327; 48-60
10. Favaudon V, Hennequin C, Targeted drugs in radiation therapy: Cancer Radiother, 2004; 8(Suppl 1); S114-20
11. Koppenhagen P, Dickreuter E, Cordes N, Head and neck cancer cell radiosensitization upon dual targeting of c-Abl and beta1-integrin: Radiother Oncol, 2017; 124; 370-78
12. Liu X, Huang W, Li C, Interaction between c-Abl and Arg tyrosine kinases and proteasome subunit PSMA7 regulates proteasome degradation: Mol Cell, 2006; 22; 317-27
13. Li D, Dong Q, Tao Q, c-Abl regulates proteasome abundance by controlling the ubiquitin-proteasomal degradation of PSMA7 subunit: Cell Rep, 2015; 10; 484-96
14. McBride WH, Iwamoto KS, Syljuasen R, The role of the ubiquitin/proteasome system in cellular responses to radiation: Oncogene, 2003; 22; 5755-73
15. Pickering AM, Davies KJ, Differential roles of proteasome and immunoproteasome regulators Pa28alphabeta, Pa28gamma and Pa200 in the degradation of oxidized proteins: Arch Biochem Biophys, 2012; 523; 181-90
16. Brooks CL, Gu W, p53 ubiquitination: Mdm2 and beyond: Mol Cell, 2006; 21; 307-15
17. Maki CG, Huibregtse JM, Howley PM: Cancer Res, 1996; 56; 2649-54
18. Bharti AC, Aggarwal BB, Nuclear factor-kappa B and cancer: Its role in prevention and therapy: Biochem Pharmacol, 2002; 64; 883-88
19. Chen ZJ, Ubiquitin signalling in the NF-kappaB pathway: Nat Cell Biol, 2005; 7; 758-65
20. Palombella VJ, Rando OJ, Goldberg AL, Maniatis T, The ubiquitin-proteasome pathway is required for processing the NF-kappa B1 precursor protein and the activation of NF-kappa B: Cell, 1994; 78; 773-85
21. Dubois C, Lecomte C, Ruys SPD: Sci Rep, 2018; 8; 11349
22. Fouad S, Wells OS, Hill MA, D’Angiolella V, Cullin ring ubiquitin ligases (CRLs) in cancer: Responses to ionizing radiation (IR) treatment: Front Physiol, 2019; 10; 1144
23. Postow MA, Callahan MK, Barker CA, Immunologic correlates of the abscopal effect in a patient with melanoma: N Engl J Med, 2012; 366; 925-31
24. Abelson HT, Rabstein LS, Influence of prednisolone on Moloney leukemogenic virus in BALB-c mice: Cancer Res, 1970; 30; 2208-12
25. Cusack JC, Rationale for the treatment of solid tumors with the proteasome inhibitor bortezomib: Cancer Treat Rev, 2003; 29(Suppl 1); 21-31
26. Cavo M, Proteasome inhibitor bortezomib for the treatment of multiple myeloma: Leukemia, 2006; 20; 1341-52
27. Brucet M, Marques L, Sebastian C, Regulation of murine Tap1 and Lmp2 genes in macrophages by interferon gamma is mediated by STAT1 and IRF-1: Genes Immun, 2004; 5; 26-35
28. Wagenblast J, Hambek M, Baghi M, Knecht R, Effect of bortezomib on EGFR expression in head and neck squamous cell carcinoma cell lines: Anticancer Res, 2008; 28; 687-92
29. Ow TJ, Thomas C, Fulcher CD, Apoptosis signaling molecules as treatment targets in head and neck squamous cell carcinoma: Laryngoscope; 2020 [Epub ahead of print]
30. Elghouche A, Shokri T, Qin Y, Unilateral cervical polyneuropathies following concurrent bortezomib, cetuximab, and radiotherapy for head and neck cancer: Case Rep Otolaryngol, 2016; 2016 2313714
31. Allen CT, Conley B, Sunwoo JB, Van Waes C: Clin Cancer Res, 2015; 21; 942-43
Figures
Figure 1. Upon stimulation with cytokines (e.g., IFN-γ), the constitutively expressed β-subunits (β1, β2, and β5) of the 20S constructive proteasome are replaced by their inducible counterparts, β1i, β2i, and β5i, respectively, which form the 20S immunoproteasome (i20S).Figure 2. (A) JHU-011 cells, FaDu cells were divided into 5 groups according to the dose gradient (0 Gy, 2.5 Gy, 5 Gy, 10 Gy, 20 Gy), and γ-irradiation was carried out at room temperature. After irradiation, the cells were cultured for 12 h, and then the RT-PCR was performed to detect the transcription level of PSMB8. The dose of 10 Gy showed the most significant induction effect on all 3 types of the cells (** P<0.01). (B) Western blot of PSMB8 in FaDu cells and JHU-011 cells. Lanes: 1, FaDu cells without irradiation; 2, FaDu cells with irradiation; 3, JHU-011 cells without irradiation; 4, JHU-011 cells with irradiation. (C) The dose of 10 Gy was used to irradiate the JHU-011 cells and FaDu cells, and the relative ratio between isoforms E1 and E2 was detected by RT-PCR. The percentage of E2 was significantly upregulated by irradiation in FaDu cells (** P<0.01) and JHU-011 cells (** P<0.01). Each group included 3 independent repeats. Quantitative data are expressed as the mean±standard deviation (SD). (D) Plasmid pLKO.1-TRC contained the insertion site of the shRNA, located between the U6 promoter and cPPT, which originally was a stuffer of approximately 1.9 kb long. After digestion with AgeI and EcoRI, the designed double-stranded shRNA oligos (c-Abl-shRNA and c-Abl-Arg) were inserted into the gap between the AgeI and EcoRI sites to replace the stuffer, yielding the vector.Figure 3. Effect of targeted silencing of the c-Abl gene on the mRNA and protein levels of PSMB8 and its alternatively spliced isoforms. (A) JHU-011 cells and FaDu cells were divided into 6 groups (described previously). In the 3 groups with IFN-γ (final concentration of 150 U/μl), the transcription of PSMB8 in the IFN-γ+/Nor group and the IFN-γ+/Arg group was significantly upregulated but was inhibited in the IFN-γ+/c-Abl-shRNA group due to the targeted silencing of the c-Abl gene (** P<0.01). (B) PSMB8 protein expression levels were detected in JHU-011 cells by Western blotting, and the ImageJ software was used to determine the gray value and drew a histogram. Lanes 1–6 represent IFN-γ−/RNAi−/Arg−, IFN-γ−/RNAi−/Arg+, IFN-γ−/RNAi+/Arg−, IFN-γ+/RNAi−/Arg−, IFN-γ+/RNAi−/Arg+, and IFN-γ+/RNAi+/Arg−, respectively. The results show that the PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg− group (** P<0.01). (C) The PSMB8 protein expression level was inhibited in the IFN-γ+/RNAi+/Arg- group of FaDu cells (** P <0.01). (D) FaDu cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group. The transcription level of isoform E2 was significantly upregulated by IFN-γ (** P<0.01). The targeted silencing of c-Abl gene (IFN-γ+/c-Abl-shRNA) downregulated the transcription of E2 (** P<0.01). (E) JHU-011 cells were divided into 6 groups. RT-PCR was performed to determine the ratio between E1 and E2 in each group, and the results were consistent with those of FaDu cells (** P<0.01). Each group contained 3 independent replicates. Quantitative data are expressed as the mean±SD.Figure 4. Regulation of tyrosine kinase activity on the transcription and expression levels of PSMB8 and its alternatively spliced isoforms (A) JHU-011 cells and FaDu cells were divided into 5 groups. In the IFN-γ+/Nilo+ group, nilotinib was first added and the cells were incubated for 24 h. Then IFN-γ was added and the cells were cultured for another 24 h. In the IFN-γ+/Nilo− group, the transcription level of PSMB8 was significantly upregulated compared to that in the control group (IFN-γ−/Nilo− group) (** P<0.01), while the transcription of PSMB8 in the IFN-γ−/Nilo+ group was inhibited (* P<0.05). (B) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated compared to that in the IFN-γ+/Nilo− group in FaDu cells (** P<0.01). (C) The protein expression of PSMB8 in the IFN-γ+/Nilo+ group was downregulated in JHU-011 cells (** P<0.01). (D) Nilotinib downregulated the induction effect of IFN-γ on isoform E2, and the percentage of isoform E2 was reduced (* P<0.05). In the DPH+ group, the transcription level of isoform E2 was higher than that in the control group (IFN-γ−/Nilo− group) (** P<0.01). Quantitative data are expressed as the mean±SD. (E) In FaDu cells, the regulation of nilotinib on the transcription of E2 was consistent with the results in JHU-011 cells (* P<0.05). (F) In the DPH+ group, protein expression of PSMB8 in JHU-011 cells and FaDu cells was elevated compared to that in the blank control group (** P<0.01). Each group contained 3 independent replicates. In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387