10 August 2020: Lab/In Vitro Research
Silencing of Long Non-Coding RNA (lncRNA) Nuclear Paraspeckle Assembly Transcript 1 (NEAT1) Protects PC-12 Cells from LPS-Induced Injury via Targeting miR-29a
Yunchao Ban1ABCD, Cui Cui2AE*DOI: 10.12659/MSM.923914
Med Sci Monit 2020; 26:e923914






Abstract
BACKGROUND: Spinal cord injury (SCI) is a debilitating neuropathological condition that significantly affects the quality of life. The present study is basic research examining the underlying mechanisms of NEAT1 and miR-29a in regulating LPS-induced PC-12 cell injury.
MATERIAL AND METHODS: The model of cell injury was induced by the treatment of PC-12 cells with LPS. The expressions of NEAT1, miR-29a, and inflammatory cytokines were measured by real-time quantitative polymerase chain reactions (RT-qPCR). Cell proliferation and apoptosis were evaluated by CCK-8 and flow cytometry, respectively. Finally, the target between miR-29a and NEAT1 as well as miR-29a and BCL2L11 was investigated by luciferase and RNA pull-down assays.
RESULTS: Knockdown of NEAT1 can inhibit inflammatory cytokine expression and PC-12 cell apoptosis and promote PC-12 cell proliferation by targeting miR-29a. However, the variation caused by NEAT1 knockdown can be reversed by the silencing of miR-29a and the overexpression of BCL2L11, which is the direct target gene of miR-29a.
CONCLUSIONS: High NEAT1 levels can increase LPS-induced injury in PC-12 cells through the miR-29a/BCL2L11 pathway. lncRNA NEAT1 may, therefore, be a promising target for SCI treatment.
Keywords: bcl-X Protein, Spinal Cord Injuries, Bcl-2-Like Protein 11, Cytokines, Gene Silencing, Gene Targeting, HEK293 Cells, Inflammation Mediators, Lipopolysaccharides
Background
Spinal cord injury (SCI), which is mainly the result of direct trauma or pathological alterations, has been identified as a critical global health priority [1]. SCI is classified as a primary or secondary injury based on pathological characteristics [2]. Primary injury is the destruction of the spinal cord structure, which contributes to the initial secondary injury [3]. Because damage to spinal cord function and structure is irreversible, the treatment strategy focuses mainly on the secondary injury [4]. However, there is still a lack of appropriate strategies or effective treatment to prevent secondary injury. There is an urgent need to broaden our understanding of the mechanism underlying the development of SCI [5]. Generally, secondary damage involves inflammation, edema, excitotoxicity, free radical production, and cell apoptosis [6]. Neuronal apoptosis is one of the first features observed in SCI, leading to various degrees of neurological damage, necrosis, or autonomic nervous dysfunction [7].
Long non-coding RNAs (lncRNA) are a class of non-coding RNAs with a length of more than 200 nucleotides that regulate cellular processes [8,9], such as cell proliferation, apoptosis, and differentiation, through the regulation of gene expression [10–12]. The aberrant expression of specific lncRNAs indicate immune system diseases [10,11]. More importantly, lncRNAs are reported to function as vital SCI regulators [15,16].
Nuclear Paraspeckle Assembly Transcript 1 (NEAT1) is a long non-coding RNA (lncRNA) primarily located in the nucleus, which is essential for the formation of nuclear paraspeckle [17]. NEAT1 plays a key role in various biological processes, including cell viability, apoptosis, DNA damage, and migration [18–21], and thus is involved in the progression of a variety of diseases. However, the effects of NEAT1 on SCI remain unknown.
This study explored the effect of NEAT1 on modulating LPS-induced injury of PC-12 cells and to elucidate the molecular mechanism. NEAT1 was found be upregulated in SCI injury and can inhibit LPS-induced apoptosis and PC-12 cell inflammatory response by controlling the miR-29a/BCL2L11 signaling axis.
Material and Methods
CELL CULTURE:
Rat pheochromocytoma adrenal gland PC-12 cells were purchased from ATCC (American Type Culture Collection, USA) and incubated in RPMI 1640 medium (Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS) with penicillin-streptomycin in a humidified atmosphere with 5% CO2 at 37°C.
VECTOR CONSTRUCTION:
Synthesized NEAT1 small interference RNA (si-NEAT1) was cloned into the vector (pAdTrack-CMV, GenePharma, Shanghai, China) and scrambled siRNA (si-nc) was cloned into the vector as a negative control. HEK-293 cells were used to pack lentivirus by transfection with vectors containing si-NEAT1 or si-nc. After 72 h of transfection, the cells were infected with virus particles using Polybrene (Sigma, St Louis, MO).
To obtain BCL2L11 overexpression plasmid, BCL2L11 was first amplified from cDNA using PCR and then inverted into pcDNA3.1 vector (Invitrogen, Carlsbad, CA) with the name of pcDNA3.1-BCL2L11.
miR-29a inhibitor, miR-29a mimics, inhibitor nc, and mimics nc were purchased from GenePharma (Shanghai, China).
After vector construction, the treated cells were transfected with si-nc, si-NEAT1, miR-29a inhibitor, inhibitor nc, miR-29a mimics, mimics nc, pcDNA3.1, or pcDNA3.1-BCL2L11 using Lipofectamine 2000 (Invitrogen, Carlsbad, CA).
REAL-TIME QUANTITATIVE POLYMERASE CHAIN REACTION (RT-QPCR) ANALYSIS:
Total RNA was extracted from cells using TRIzol reagent (Thermo Fisher Scientific., Inc, Waltham, MA, USA) following the manufacturer’s instructions. Then, cDNA was synthesized from RNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA). Then, quantitative real-time PCR was carried out using SYBR Green PCR Master Mix with a Fast Real-time PCR 7500 System (Applied Biosystem, Carlsbad, CA, USA). GAPDH and U6 functioned as internal controls. Data were analyzed according to the 2−ΔΔCt method and normalized to internal controls.
WESTERN BLOT ANALYSIS:
After transfection, PC-12 cells were lysed in Triton X-100 lysis buffer and protease inhibitor cocktail (Sigma-Aldrich, Shanghai, China) at 4°C. The concentrations of proteins were measured using the BCA assay kit (Pierce, Appleton, WI, USA). Then, the proteins were separated and transferred from 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE; SolarBio Life Sciences, Beijing, China) to polyvinylidene fluoride (PVDF; SolarBio Life Sciences, Beijing, China) membranes. Afterward, the membrane was blocked in non-fat milk for 50 min and incubated with primary antibodies overnight at 4°C. Finally, horseradish peroxidase-conjugated anti-rabbit IgG secondary antibodies and enhanced Luminata Classic Western HRP Substrate were added and the blots were visualized using ImageQuant LAS 4000 mini (GE Healthcare, NJ, USA).
ELISA ASSAY:
Cell supernatants were collected and determined using ELISA kits (Thermo Fisher Scientific., Inc, Waltham, MA, USA) according to the manufacturer’s protocol. The optical density was detected at 450 nm, and the results were calculated using the linear calibration curves generated by standard solutions.
CELL VIABILITY ANALYSIS:
After transfection and LPS treatment, the cells were grown in 96-well-plated cells with a density of 1×103 cells/well. At different time points, the Cell Count Kit-8 (CCK-8; Beyotime, Shanghai, China) was added to each well and incubated for another 2 h at 37°C. The absorbance value of each well was detected at 450 nm using a microplate reader (Bio-Tek Instruments, Winooski, VT, USA) in triplicate.
CELL APOPTOSIS ANALYSIS:
Cell apoptosis was detected using an Annexin V-FITC apoptosis detection kit (Beyotime Biotechnology, Shanghai, China) and analyzed with BD FACSDiva software version 6.1.3 (BD Bioscience, USA). Briefly, after transfection and LPS treatment, cells were grown in a 6-well plate with a density of 1×105 cells/well in a humidified atmosphere with 5% CO2 for 48 h. The cells were then washed with ice-cold PBS and stained with Annexin V-FITC and PI according to the manufacturer’s instructions. All of the above procedures were carried out in triplicate.
LUCIFERASE REPORTER ASSAY:
NEAT1-mutant and BCL2L11-mutant plasmids were designed and purchased from GeneScript (Nanjing, China). The 3′-UTR of lncRNA NEAT1 and BCL2L11 cRNA fragments containing the potential binding sequences of miR-29a sites were amplified from PCR and inverted into pGL3 vector (Ambion, Inc., Austin, TX, USA).
HEK-293 cells were cultured in 24-well plates and co-transfected with vectors and miR-29a mimics or miR-nc. Afterward, cells were transfected using Lipofectamine 2000 (Thermo Fisher Scientific., Inc, Waltham, MA, USA). Finally, the luciferase activities were detected using a dual-luciferase reporter assay system (Promega, Madison, WI) following the manufacturer’s instructions.
BIOTIN-LABELED MIR-29A PULL-DOWN ASSAY:
Biotin-labeled negative control (biotin-nc) or biotin-labeled miR-29a (biotin-miR-29a) were transfected into treated HEK-293 cells. Afterward, cell lysate was collected and mixed with streptavidin-coupled Dynabeads (Invitrogen, Carlsbad, CA) at 4°C overnight on a rotator. Finally, the bead-bound RNA was washed and isolated, followed by RT-qPCR analysis. Input RNA was extracted and functioned as controls.
STATISTICAL ANALYSIS:
SPSS 18.0 (IBM SPSS Statistics, USA) and GraphPad Prism software 5.0 (GraphPad Prism Software Inc., San Diego, CA) were used to analyze the data. All data are presented as mean±SD (standard deviation). The
Results
LPS INHIBITED THE PROLIFERATION AND INDUCED APOPTOSIS AND INFLAMMATORY RESPONSE OF PC-12 CELL:
First, we established the LPS-induced PC-12 cell injury model. As Figure 1 shows, LPS of 5, 10, and 20 μg/ml significantly inhibited the proliferation of PC-12 cells (Figure 1A) and notably promoted apoptosis (Figure 1B). We further evaluated the expression of apoptotic proteins. LPS significantly promoted the expression of pro-apoptosis proteins such as bax and cleaved caspase and inhibited that of anti-apoptosis protein bcl-2 (Figure 1C). ELISA was used to assess the expression of inflammatory factors; the results indicated that expression of inflammatory factors including IL-1β, IL-6, and TNF-α of PC-12 cells was promoted by LPS treatment, while expression of IL-10 was inhibited (Figure 1D). These results indicated that we successfully established the LPS-induced cell injury model. Then, we evaluated the expression of NEAT1 in PC-12 cells treated by LPS. NEAT1 was significantly increased in PC-12 cells after treatment with 5, 10, and 20 μg/ml of LPS (Figure 1E).
KNOCKDOWN OF NEAT1 REVERSED THE EFFECT OF LPS ON PROLIFERATION, APOPTOSIS, AND INFLAMMATORY RESPONSE OF PC-12 CELLS:
As NEAT1 was notably upregulated in LPS-treated PC-12 cells. We speculated that NEAT1 plays a critical role in LPS-induced injury of PC-12 cells. The lentivirus knocking down NEAT1 was obtained. As demonstrated in Figure 2A, the expression of NEAT1 was markedly decreased in the si-NEAT1 group compared to si-nc and control groups. The CCK-8 assay further demonstrated that after LPS treatment, the proliferation of PC-12 cells was significantly decreased; however, the decreased viability could be partially reversed by transfection with Ad-siNEAT1 (Figure 2B). LPS promoted the expression of inflammatory factors such as IL-1β, IL-6, and TNF-α and inhibited that of IL-10. Infection with si-NEAT1 reversed the LPS-induced changes in inflammatory cytokines expressions (Figure 2C). Moreover, the PC-12 cell apoptosis was increased after LPS treatment and this was partially knocked down by Ad-siNEAT1 infection (Figure 2D, 2E).
RECIPROCAL INHIBITION BETWEEN LNCRNA NEAT1 AND MIR-29A IN HEK-293 CELLS:
We used the bioinformatics website Starbase to predict the potential targets for lncRNA NEAT1. As shown in Figure 3A, the binding sites of miR-29a were found within the sequence of NEAT1. Afterward, the luciferase reporter assay confirmed that the co-transfection of HEK-293 cells with wild-type lncRNA NEAT1 and the miR-29a mimics resulted in remarkably decreased luciferase activity as compared with the miR-control group (Figure 3B). Then, the RNA pull-down assay verified that the biotinylated miR-29a enriched the expression of NEAT1, which verified the interaction between NEAT1 and miR-20a (Figure 3C). Meanwhile, the relative expression of miR-29a was remarkably increased in the si-NEAT1 group compared with the si-nc group (Figure 3D). Collectively, these results showed that miR-29a was a direct target of lncRNA NEAT1.
KNOCKDOWN OF MIR-29A PARTIALLY REVERSED THE EFFECT OF NEAT1 ON THE PROLIFERATION, APOPTOSIS, AND INFLAMMATORY RESPONSE:
As demonstrated in Figure 4A, infection with Ad-si-NEAT1 remarkably increased the expression of miR-29a after LPS treatment; however, the increase was counteracted by co-infection with Ad-si-miR-29a. The CCK-8 assay further demonstrated that after LPS treatment, the proliferation of PC-12 cells was significantly decreased. After infection with Ad-si-NEAT1, cell viability was remarkably increased; however, the increase was partially counteracted by co-infection with Ad-si-miR-29a (Figure 4B). After LPS treatment, the apoptosis of PC-12 cells was significantly increased. After infection with Ad-si-NEAT1, cell apoptosis was remarkably decreased; however, the decrease was partially reversed by co-infection with Ad-si-miR-29a (Figure 4C–4E). Similarly, the expression levels of IL-1β, IL-6, and TNF-α in PC-12 cells were significantly increased, while IL-10 expression was lower in the LPS group compared with the untreated group. After co-transfection with Ad-siNEAT1 and Ad-si-nc, expression levels of IL-1β, IL-6, and TNF-α were significantly inhibited, while IL-10 expression was increased. However, transfection with Ad-si-NEAT1 and Ad-si-miR-29a inhibitor counteracted the Ad-siNEAT1-induced changes in inflammatory cytokines expressions (Figure 4F).
MIR-29A TARGETS BCL2L11 GENE IN HEK-293 CELLS:
The bioinformatics website TargetScan was used to predict the potential targets for miR-29a. As shown in Figure 5A, the binding sites of BCL2L11 were found within miR-29a miRNA. Afterward, the dual-luciferase reporter assay confirmed that the co-transfection of HEK-293 cells with wild-type BCL2L11 3′-UTR and the miR-29a mimics resulted in remarkably decreased luciferase activity as compared with the miR-nc group (Figure 5B). Meanwhile, relative mRNA and protein expressions of BCL2L11 were remarkably decreased in the miR-29a mimics group compared with the miR-nc group; however, BCL2L11 expressions were remarkably increased after transfected with miR-29a inhibitor (Figure 5C, 5D). Then, the RNA pull-down assay verified that the biotinylated miR-29a enriched the expression of BCL2L11, which confirmed the interaction between miR-29a and BCL2L11 (Figure 5E). The correlation results in Figure 5F demonstrated that BCL2L11 expression was negatively correlated with miR-29a (r2=0.3335; p=0.00242). Collectively, these results showed that BCL2L11 was a direct target of miR-29a.
NEAT1 ACTS AS THE CERNA OF BCL2L11 AND REGULATES APOPTOTIC PROTEIN EXPRESSION:
Pearson correlation analysis demonstrated that BCL2L11 expression was positively correlated with NEAT1 (r2=0.2387; p=0.0132) (Figure 6A), indicating that NEAT1 functions as a ceRNA of BCL2L11 in PC-12 cells. Moreover, Western blot results revealed that LPS promoted the expression of BCL2L11, bax, and cleaved caspase3 and inhibited that of bcl-2. However, infection of Ad-si-NEAT1 notably reduced the level of BCL2L11, bax, and cleaved caspase3 and increased that of bcl-2 compared to the LPS group. Transfection of BCL2L11-overexpressing vector partially counteracted the effect of Ad-si-NEAT1, which further verified the ceRNA correlation between NEAT1 and BCL2L11 (Figure 6B).
Discussion
The present study explored the interplay between lncRNA NEAT1 and miR-29a and its target gene BCL2L11 in regulating PC-12 cellular processes after LPS treatment.
Recently, accumulating reports suggested that some lncRNAs and miRNAs are involved in the pathogenesis and development of spinal cord injury [22–24]. Our results indicated that lncRNA NEAT1 was significantly increased while miR-29a was decreased in PC-12 cells treated with LPS. Moreover, downregulation of lncRNA NEAT1 promoted LPS-induced PC-12 cell proliferation but inhibited apoptosis; however, this was reversed by co-transfection with miR-29a.
The role of NEAT1 in spinal cord injury has been unclear. Previous studies have shown that Neat1 is upregulated in the spinal cord tissues of chronic constriction injury rats. Neat1 downregulation suppresses neuropathic pain and neuroinflammation in CCI rats [25]. We found a similar effect of NEAT1 in LPS-induced PC-12 cell injury. Another study indicated that Neat1 promotes neuronal differentiation and migration and inhibits apoptosis of spinal cord neural stem cells (SC-NPCs). Moreover, it can be upregulated by miR-124 and is involved in activating Wnt/β-catenin signaling. We speculate that NEAT1 exerts distinct roles in different cells, such as PC-12 and SC-NPSs. Previous studies did not sufficiently elucidate the mechanism underlying the effect of NEAT1. The common regulating mode of miRs is well-established, but how miR-124 upregulates the expression of NEAT1 remains unknown. In our study, a luciferase reporter assay, RNA pull-down, and Pearson analysis confirmed that miR-29a was a direct target of LncRNA NEAT1 and could directly inhibit its downstream protein BCL2L11. BCL2L11 is well known to be a pro-apoptosis protein [26]. NEAT1 acts as a ceRNA of BCL2L11 to participate in the regulation of apoptosis.
miR-29a has been widely studied in the central nervous system [27]. miR-29a can promote neuronal differentiation but inhibits the astrocytes differentiation of neural stem cells by downregulating PTEN expression [28]. It can also inhibit the apoptosis of hippocampal neurons induced by oxygen and glucose deprivation/reperfusion (OGD/R) [29]. Interestingly, we found that miR-29a modulates neurite outgrowth in PC-12 cells. miR-29a directly targets PTEN and modulates the phosphorylation of AKT [30]. Overexpression of miR-29a mediated by lentivirus can promote functional recovery in spinal cord injury by promoting axonal regeneration [31]. These findings indicate the protective roles of miR-29a. We confirmed that miR-29a could protect the PC-12 cells from LPS-induced apoptosis and inflammation response via directly targeting BCL2L11. The present results extend our understanding of the role and mechanism of miR-29a in spinal cord injury.
Inflammation is associated with the pathogenesis and development of SCI [32,33]. Our results showed that IL-1β, IL-6, and TNF-α expressions were decreased by NEAT1 knockdown while IL-10 expression was increased. This effect of NEAT1 can be partially reversed by miR-29a. However, the precise mechanism by which NEAT1 and miR-29a regulate the inflammatory response remain unknown, and
Conclusions
Our findings show the effect of NEAT1 on proliferation, apoptotic, and inflammatory response of PC-12 cells treated with LPS. NEAT1 exerts a protective role against LPS-induced cell injury via miR-29a/BCL2L11 signaling. Our findings suggest that NEAT1 is a potential indicator for the treatment of SCI.
Figures
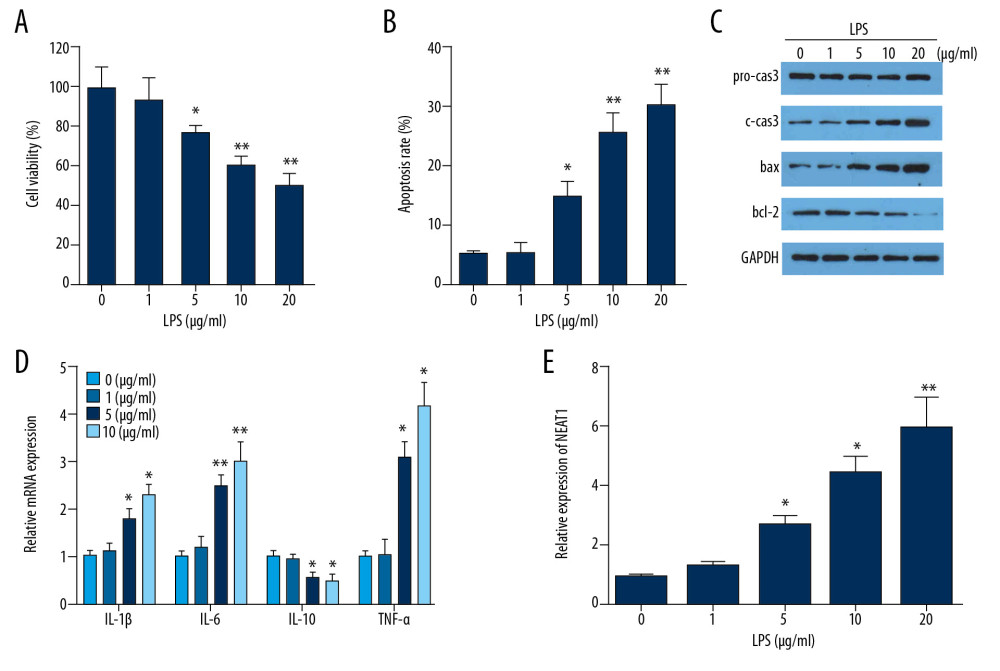 Figure 1. The effect of LPS treatment on PC-12 cells. (A) PC-12 cell viability after LPS treatment was detected by CCK-8. (B) PC-12 cell apoptosis after LPS treatment was detected by flow cytometry. (C, D) Expression levels of inflammatory cytokines after LPS treatment were detected by ELISA. (E) Relative expression of lncRNA NEAT1 after LPS treatment was assessed by qPCR. * P<0.05 vs. 0 μg/ml, ** P<0.01 vs. 0 μg/ml.
Figure 1. The effect of LPS treatment on PC-12 cells. (A) PC-12 cell viability after LPS treatment was detected by CCK-8. (B) PC-12 cell apoptosis after LPS treatment was detected by flow cytometry. (C, D) Expression levels of inflammatory cytokines after LPS treatment were detected by ELISA. (E) Relative expression of lncRNA NEAT1 after LPS treatment was assessed by qPCR. * P<0.05 vs. 0 μg/ml, ** P<0.01 vs. 0 μg/ml. 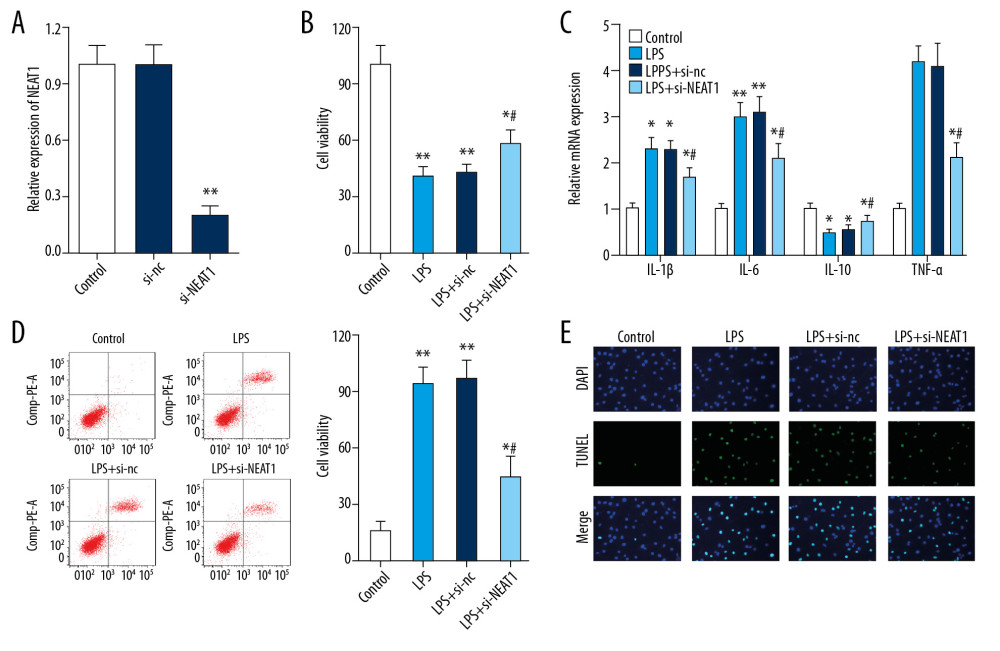 Figure 2. The effect of NEAT1 knockdown on PC-12 cells after LPS treatment. (A) The relative expression of LncRNA NEAT1 after Ad-si-NEAT1 infection. (B) PC-12 cell viability after LPS treatment and Ad-si-NEAT1 infection was detected by CCK-8. (C) Expression levels of inflammatory cytokines after LPS treatment were evaluated by ELISA. (D, E) PC-12 cell apoptosis after LPS treatment and Ad-si-NEAT1 infection was detected by flow cytometry and TUNEL staining. * P<0.05 vs. control,** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc.
Figure 2. The effect of NEAT1 knockdown on PC-12 cells after LPS treatment. (A) The relative expression of LncRNA NEAT1 after Ad-si-NEAT1 infection. (B) PC-12 cell viability after LPS treatment and Ad-si-NEAT1 infection was detected by CCK-8. (C) Expression levels of inflammatory cytokines after LPS treatment were evaluated by ELISA. (D, E) PC-12 cell apoptosis after LPS treatment and Ad-si-NEAT1 infection was detected by flow cytometry and TUNEL staining. * P<0.05 vs. control,** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc. 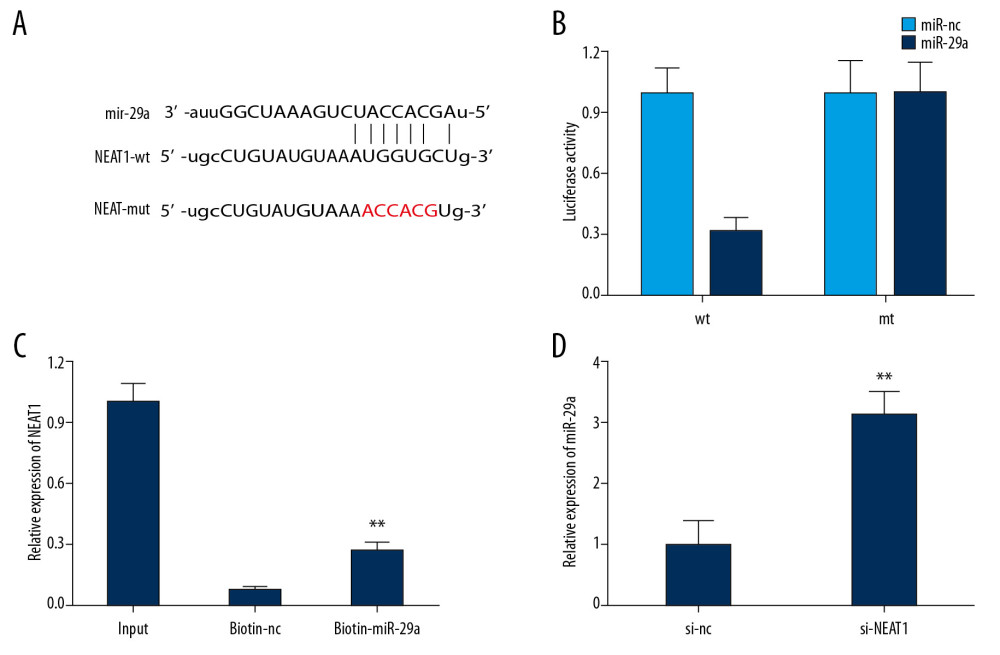 Figure 3. Reciprocal inhibition between lncRNA NEAT1 and miR-29a in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within lncRNA NEAT1 mRNA. (B) The relative luciferase activities were detected by dual-luciferase reporter assay in HEK-293 cells co-transfected with wild-type and mutant-type vectors together with miR-29a mimics and miR-nc. (C) The expression level of lncRNA NEAT1 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (D) The relative expression of miR-29a in PC-12 cells after infection with Ad-si-NEAT1 or si-nc. * P<0.05 vs. miR-nc, ** P<0.01 vs. biotin-nc or si-nc.
Figure 3. Reciprocal inhibition between lncRNA NEAT1 and miR-29a in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within lncRNA NEAT1 mRNA. (B) The relative luciferase activities were detected by dual-luciferase reporter assay in HEK-293 cells co-transfected with wild-type and mutant-type vectors together with miR-29a mimics and miR-nc. (C) The expression level of lncRNA NEAT1 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (D) The relative expression of miR-29a in PC-12 cells after infection with Ad-si-NEAT1 or si-nc. * P<0.05 vs. miR-nc, ** P<0.01 vs. biotin-nc or si-nc. 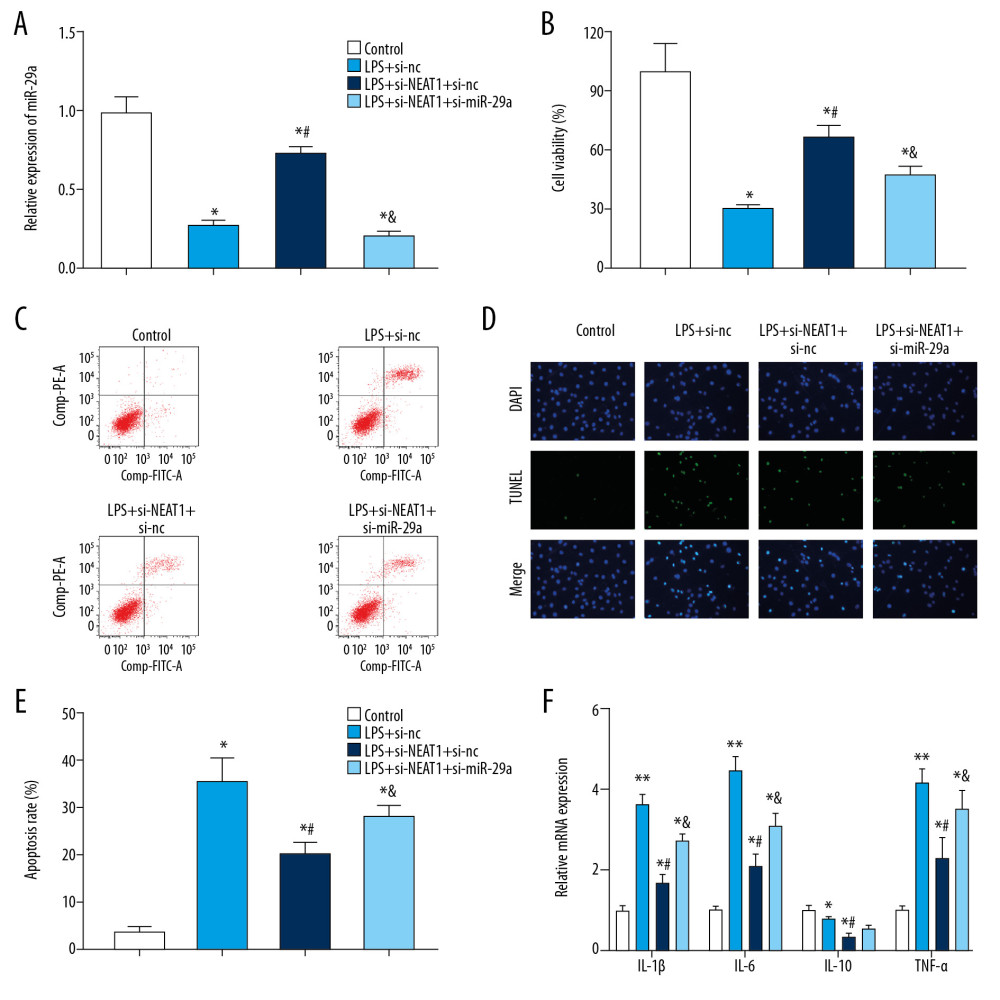 Figure 4. Upregulation of miR-29a counteracted the changes induced by Ad-si-NEAT1 in PC-12 cells after LPS treatment. (A) The relative expression of miR-29a in different groups was evaluated by qPCR. (B) PC-12 cell viability after LPS treatment or transduction of Ad-si-NEAT1 or si-miR-29a was assessed by CCK-8. (C–E) PC-12 cell apoptosis was evaluated by flow cytometry and TUNEL staining. (F) Expression levels of inflammatory cytokines was detected by ELISA assay. * P<0.05 vs. control, ** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc, & P<0.05 vs. LPS+si-NEAT1+si-nc.
Figure 4. Upregulation of miR-29a counteracted the changes induced by Ad-si-NEAT1 in PC-12 cells after LPS treatment. (A) The relative expression of miR-29a in different groups was evaluated by qPCR. (B) PC-12 cell viability after LPS treatment or transduction of Ad-si-NEAT1 or si-miR-29a was assessed by CCK-8. (C–E) PC-12 cell apoptosis was evaluated by flow cytometry and TUNEL staining. (F) Expression levels of inflammatory cytokines was detected by ELISA assay. * P<0.05 vs. control, ** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc, & P<0.05 vs. LPS+si-NEAT1+si-nc. 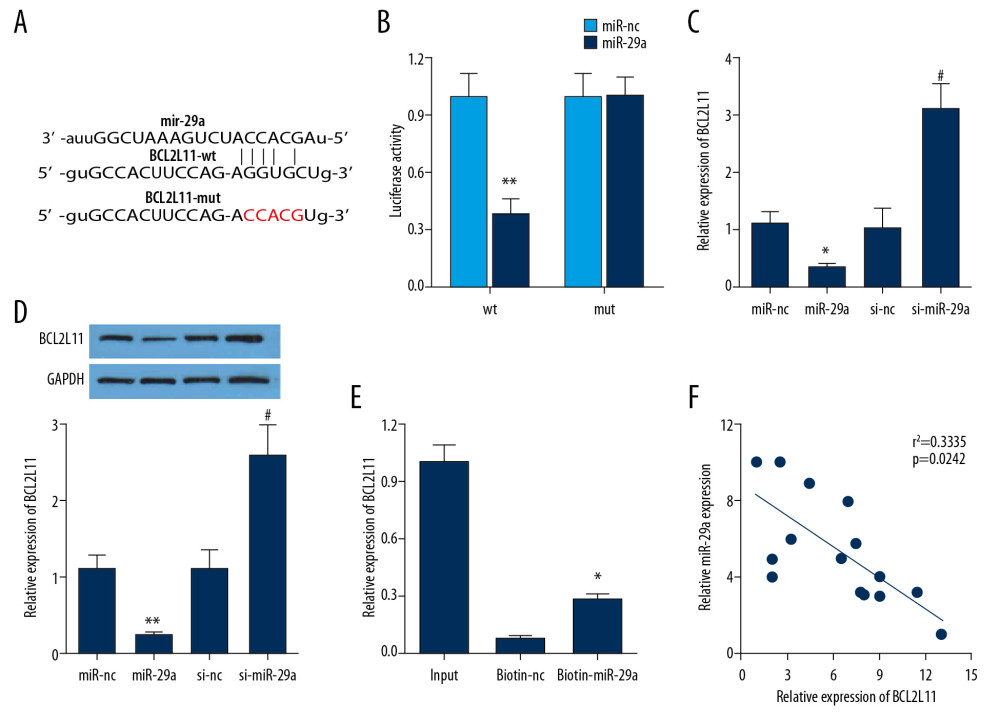 Figure 5. miR-29a targeted the BCL2L11 gene in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within BCL2L11 mRNA. (B) The relative luciferase activities were detected in HEK-293 cells co-transfected with vectors containing the wild- or mutant-type of BCL2L11 together with miR-29a mimics or miR-nc. (C, D) The relative mRNA and protein expression of BCL2L11 after transfection of miR-29a mimic or si-miR-29a. (E) The expression level of BCL2L11 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (F) The correlation between miR-29a and BCL2L11 was analyzed by Pearson analysis. * P<0.05 vs. miR-nc or biotin-nc, ** P<0.01 vs. miR-nc, # P<0.01 vs. si-nc.
Figure 5. miR-29a targeted the BCL2L11 gene in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within BCL2L11 mRNA. (B) The relative luciferase activities were detected in HEK-293 cells co-transfected with vectors containing the wild- or mutant-type of BCL2L11 together with miR-29a mimics or miR-nc. (C, D) The relative mRNA and protein expression of BCL2L11 after transfection of miR-29a mimic or si-miR-29a. (E) The expression level of BCL2L11 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (F) The correlation between miR-29a and BCL2L11 was analyzed by Pearson analysis. * P<0.05 vs. miR-nc or biotin-nc, ** P<0.01 vs. miR-nc, # P<0.01 vs. si-nc. 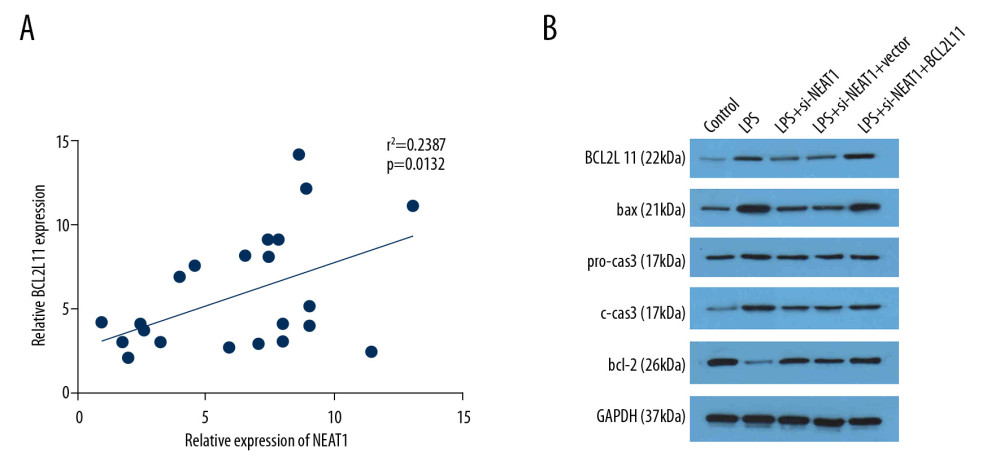 Figure 6. NEAT1 modulated the BCL2L11 and the apoptotic signal pathway. (A) Pearson analysis was performed to evaluate the correlation between NEAT1 and BCL2L11. (B) The expression of BCL2L11, bax, pro-caspase3, caspase3, and bcl-2 was evaluated by Western blot.
Figure 6. NEAT1 modulated the BCL2L11 and the apoptotic signal pathway. (A) Pearson analysis was performed to evaluate the correlation between NEAT1 and BCL2L11. (B) The expression of BCL2L11, bax, pro-caspase3, caspase3, and bcl-2 was evaluated by Western blot. References
1. Cadel L, Everall AC, Packer TL, Exploring the perspectives on medication self-management among persons with spinal cord injury/dysfunction and providers: Res Social Adm Pharm, 2020 [Online ahead of print]
2. James ND, McMahon SB, Field-Fote EC, Bradbury EJ, Neuromodulation in the restoration of function after spinal cord injury: Lancet Neurol, 2018; 17; 905-17
3. Fogarty MJ, Sieck GC, Spinal cord injury and diaphragm neuromotor control: Expert Rev Respir Med, 2020; 14; 553-64
4. de Araujo A, Ribeiro F, Massetti T, Effectiveness of anodal transcranial direct current stimulation to improve muscle strength and motor functionality after incomplete spinal cord injury: a systematic review and meta-analysis: Spinal Cord, 2020 [Online ahead of print]
5. Ong B, Wilson JR, Henzel MK, Management of the patient with chronic spinal cord injury: Med Clin North Am, 2020; 104; 263-78
6. Kim HY, Kumar H, Jo MJ, Therapeutic efficacy-potentiated and diseased organ-targeting nanovesicles derived from mesenchymal stem cells for spinal cord injury treatment: Nano Lett, 2018; 18; 4965-75
7. Ramer LM, Ramer MS, Bradbury EJ, Restoring function after spinal cord injury: Towards clinical translation of experimental strategies: Lancet Neurol, 2014; 13; 1241-56
8. Shi X, Sun M, Liu H, Long non-coding RNAs: A new frontier in the study of human diseases: Cancer Lett, 2013; 339; 159-66
9. Dey BK, Mueller AC, Dutta A, Long non-coding RNAs as emerging regulators of differentiation, development, and disease: Transcription, 2014; 5; e944014
10. Vadaie N, Morris KV, Long antisense non-coding RNAs and the epigenetic regulation of gene expression: Biomol Concepts, 2013; 4; 411-15
11. Wang JZ, Xu CL, Wu H, Shen SJ, LncRNA SNHG12 promotes cell growth and inhibits cell apoptosis in colorectal cancer cells: Braz J Med Biol Res, 2017; 50; e6079
12. Jiang W, Liu Y, Liu R, The lncRNA DEANR1 facilitates human endoderm differentiation by activating FOXA2 expression: Cell Rep, 2015; 11; 137-48
13. Atianand MK, Fitzgerald KA, Long non-coding RNAs and control of gene expression in the immune system: Trends Mol Med, 2014; 20; 623-31
14. Stachurska A, Zorro MM, van der Sijde MR, Withoff S, Small and long regulatory RNAs in the immune system and immune diseases: Front Immunol, 2014; 5; 513
15. Yu TM, Palanisamy K, Sun KT, RANTES mediates kidney ischemia reperfusion injury through a possible role of HIF-1alpha and LncRNA PRINS: Sci Rep, 2016; 6; 18424
16. Zhou P, Chen Z, Zou Y, Wan X, Roles of non-coding RNAs in acute kidney injury: Kidney Blood Press Res, 2016; 41; 757-69
17. Machitani M, Taniguchi I, Ohno M, ARS2 regulates nuclear paraspeckle formation through 3′-End processing and stability of NEAT1 long noncoding RNA: Mol Cell Biol, 2020; 40; e00269-19
18. Boros FA, Maszlag-Torok R, Vecsei L, Klivenyi P, Increased level of NEAT1 long non-coding RNA is detectable in peripheral blood cells of patients with Parkinson’s disease: Brain Res, 2020; 1730 146672
19. Wan Y, Yang ZQ, LncRNA NEAT1 affects inflammatory response by targeting miR-129-5p and regulating Notch signaling pathway in epilepsy: Cell Cycle, 2020; 19; 419-31
20. Liu D, Zou Z, Li G, Pan P, Liang G, Long noncoding RNA NEAT1 suppresses proliferation and promotes apoptosis of glioma cells via downregulating MiR-92b: Cancer Control, 2020; 27 1147333241
21. Adriaens C, Rambow F, Bervoets G, The long noncoding RNA NEAT1_1 is seemingly dispensable for normal tissue homeostasis and cancer cell growth: RNA, 2019; 25; 1681-95
22. Brandenburger T, Salgado SA, Devaux Y, Lorenzen JM, Noncoding RNAs in acute kidney injury: Kidney Int, 2018; 94; 870-81
23. Liu Z, Wang S, Mi QS, Dong Z, MicroRNAs in pathogenesis of acute kidney injury: Nephron, 2016; 134; 149-53
24. Fan PC, Chen CC, Chen YC, MicroRNAs in acute kidney injury: Hum Genomics, 2016; 10; 29
25. Xia LX, Ke C, Lu JM, NEAT1 contributes to neuropathic pain development through targeting miR-381/HMGB1 axis in CCI rat models: J Cell Physiol, 2018; 233; 7103-11
26. Hutcheson J, Perlman H, BH3-only proteins in rheumatoid arthritis: Potential targets for therapeutic intervention: Oncogene, 2008; 27(Suppl 1); S168-75
27. Gao Y, Qiao H, Lu Z, Hou Y, miR29 promotes the proliferation of cultured rat neural stem/progenitor cells via the PTEN/AKT signaling pathway: Mol Med Rep, 2019; 20; 2111-18
28. Shi Z, Zhou H, Lu L, MicroRNA-29a regulates neural stem cell neuronal differentiation by targeting PTEN: J Cell Biochem, 2018; 119; 5813-20
29. Wei R, Zhang R, Li H, MiR-29 Targets PUMA to suppress oxygen and glucose deprivation/reperfusion (OGD/R)-induced cell death in hippocampal neurons: Curr Neurovasc Res, 2018; 15; 47-54
30. Zou H, Ding Y, Wang K, MicroRNA-29A/PTEN pathway modulates neurite outgrowth in PC12 cells: Neuroscience, 2015; 291; 289-300
31. Yin H, Shen L, Xu C, Liu J, Lentivirus-mediated overexpression of miR-29a promotes axonal regeneration and functional recovery in experimental spinal cord injury via PI3K/Akt/mTOR pathway: Neurochem Res, 2018; 43; 2038-46
32. Sato Y, Yanagita M, Immune cells and inflammation in AKI to CKD progression: Am J Physiol Renal Physiol, 2018; 315; F1501-12
33. Cunha MI, Su M, Cantuti-Castelvetri L, Pro-inflammatory activation following demyelination is required for myelin clearance and oligodendrogenesis: J Exp Med, 2020; 217; e20191390
Figures
Figure 1. The effect of LPS treatment on PC-12 cells. (A) PC-12 cell viability after LPS treatment was detected by CCK-8. (B) PC-12 cell apoptosis after LPS treatment was detected by flow cytometry. (C, D) Expression levels of inflammatory cytokines after LPS treatment were detected by ELISA. (E) Relative expression of lncRNA NEAT1 after LPS treatment was assessed by qPCR. * P<0.05 vs. 0 μg/ml, ** P<0.01 vs. 0 μg/ml.Figure 2. The effect of NEAT1 knockdown on PC-12 cells after LPS treatment. (A) The relative expression of LncRNA NEAT1 after Ad-si-NEAT1 infection. (B) PC-12 cell viability after LPS treatment and Ad-si-NEAT1 infection was detected by CCK-8. (C) Expression levels of inflammatory cytokines after LPS treatment were evaluated by ELISA. (D, E) PC-12 cell apoptosis after LPS treatment and Ad-si-NEAT1 infection was detected by flow cytometry and TUNEL staining. * P<0.05 vs. control,** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc.Figure 3. Reciprocal inhibition between lncRNA NEAT1 and miR-29a in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within lncRNA NEAT1 mRNA. (B) The relative luciferase activities were detected by dual-luciferase reporter assay in HEK-293 cells co-transfected with wild-type and mutant-type vectors together with miR-29a mimics and miR-nc. (C) The expression level of lncRNA NEAT1 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (D) The relative expression of miR-29a in PC-12 cells after infection with Ad-si-NEAT1 or si-nc. * P<0.05 vs. miR-nc, ** P<0.01 vs. biotin-nc or si-nc.Figure 4. Upregulation of miR-29a counteracted the changes induced by Ad-si-NEAT1 in PC-12 cells after LPS treatment. (A) The relative expression of miR-29a in different groups was evaluated by qPCR. (B) PC-12 cell viability after LPS treatment or transduction of Ad-si-NEAT1 or si-miR-29a was assessed by CCK-8. (C–E) PC-12 cell apoptosis was evaluated by flow cytometry and TUNEL staining. (F) Expression levels of inflammatory cytokines was detected by ELISA assay. * P<0.05 vs. control, ** P<0.01 vs. control, # P<0.05 vs. LPS+si-nc, & P<0.05 vs. LPS+si-NEAT1+si-nc.Figure 5. miR-29a targeted the BCL2L11 gene in HEK-293 cells. (A) The sequences of wild-type and mutant-type vectors and putative binding sites of miR-29a within BCL2L11 mRNA. (B) The relative luciferase activities were detected in HEK-293 cells co-transfected with vectors containing the wild- or mutant-type of BCL2L11 together with miR-29a mimics or miR-nc. (C, D) The relative mRNA and protein expression of BCL2L11 after transfection of miR-29a mimic or si-miR-29a. (E) The expression level of BCL2L11 mRNA was measured by RT-qPCR in the sample pulled down by biotinylated miR-29a. (F) The correlation between miR-29a and BCL2L11 was analyzed by Pearson analysis. * P<0.05 vs. miR-nc or biotin-nc, ** P<0.01 vs. miR-nc, # P<0.01 vs. si-nc.Figure 6. NEAT1 modulated the BCL2L11 and the apoptotic signal pathway. (A) Pearson analysis was performed to evaluate the correlation between NEAT1 and BCL2L11. (B) The expression of BCL2L11, bax, pro-caspase3, caspase3, and bcl-2 was evaluated by Western blot. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387