02 September 2020: Clinical Research
miR-22-3p Suppresses Endothelial Progenitor Cell Proliferation and Migration via Inhibiting Onecut 1 (OC1)/Vascular Endothelial Growth Factor A (VEGFA) Signaling Pathway and Its Clinical Significance in Venous Thrombosis
Huoqi Liang1ABDE, Yibiao Chen1BCDF, Hefei Li1CDF, Xiaoqiang Yu1CEF, Chunqiu Xia1CF, Zhibing Ming1AEG*, Chongjun Zhong1AFGDOI: 10.12659/MSM.925482
Med Sci Monit 2020; 26:e925482






Abstract
BACKGROUND: Proliferation and migration play crucial roles in various physiological processes, especially in injured endothelial repair. Endothelial progenitor cells (EPCs), as the precursors of endothelial cell, are involved in the regeneration of the endothelial lining of blood vessels. Furthermore, EPCs were found to be a potential choice for venous thrombosis (VT) treatment.
MATERIAL AND METHODS: EPCs were isolated from human peripheral blood of healthy adults and VT patients. Differently expressed micro(mi)RNAs were examined by quantitative real-time polymerase chain reaction, after which proliferative capacity and migration effect were tested by Cell-Counting Kit 8, scratch wound assay, and transwell assays. Bioinformatic analysis was applied to investigate the potential target messenger ribonucleic acid and a dual-luciferase reporting system was utilized to confirm the binding of miR-22-3p to its target gene. Western blot was carried out to detect candidate protein expression level. Finally, miR-22-3p expression was monitored in VT patients during follow-up to assess its correlation with prognosis of VT.
RESULTS: Our data revealed that miR-22-3p was upregulated in EPCs derived from deep VT (DVT) individuals and suppression of miR-22-3p contributed to proliferation and migration of EPCs. In addition, miR-22-3p/onecut 1 (OC1)/vascular endothelial growth factor A (VEGFA) signaling pathway was involved in regulating EPC migration and proliferation. In addition, lower expression of miR-22-3p in DVT patients indicated decreased risk of VT recurrence.
CONCLUSIONS: Our results suggest that miR-22-3p regulates OC1/VEGFA signaling and is involved in regulating EPC proliferation and migration. The expression level of miR-22-3p could be monitored to predict DVT patients’ prognosis.
Keywords: Venous Thrombosis, Case-Control Studies, Onecut Transcription Factors
Background
Endothelial progenitor cells (EPCs) are likely important in wound healing and tissue repair for their ability to develop into mature endothelial cells [1]. Circulating EPCs were confirmed in the bloodstream of cardiovascular patients, which indicated their potential role in cardiovascular diseases. Several studies have reported that larger amount of EPCs predicted better outcome in patients with heart attack history on the basis of the observation that EPCs were mobilized after a myocardial infarction and participated in restoring the lining of blood vessels that were impaired during a heart attack [2]. However, few studies have focused on the function of EPCs in venous thrombosis (VT). Deep VT (DVT) refers the formation of a blood clot in a lower extremity. The traditional treatment regimen for DVT includes anticoagulants and thrombolytics, both of which have few benefits for injured endothelium or for the vessels. On the basis of the knowledge of the biological characteristics of EPCs, an emerging concept regarding the potential application of EPCs for therapeutic use may offer a new strategy for the treatment of VT. Herein, we aim to further elucidate the role of EPCs in the setting of DVT.
Microribonucleic acids (miRNAs), with lengths of 20–24 nucleotides, have been proposed to have the capacity to function in RNA splicing and posttranscriptional regulation of gene expression [3]. Accumulating evidence suggests that miRNAs are widely involved in the normal functioning of cells, and dysregulation of miRNA is associated with disease [4,5]. Advanced studies showed that miRNAs, including miR-126, let-7e-5p, and miR-120, participated in biological functioning of EPCs [6–8]. MiR-22-3p has been identified as an inhibitor of arterial smooth muscle cell proliferation and migration in arteriosclerosis obliterans [9]. In addition, previous studies identified miR-22-3p in peripheral blood as a biomarker for certain diseases [10]. However, the expression level and specific role of miR-22-3p in EPCs remains unclear.
In this study, we investigated the molecular function of miR-22-3p as well as its regulatory mechanisms in EPCs. Our results revealed that EPCs are regulated by miR-22-3p via regulation of onecut 1 (OC1)/vascular endothelial growth factor A (VEGFA) signaling. Notably, we identified miR-22-3p as a biomarker for the prognosis of DVT.
Material and Methods
CELL CULTURE AND TRANSFECTION:
Fifty-milliliter peripheral blood samples were collected from healthy adults and DVT patients. Informed consent was obtained from all participants and the protocol was approved by the Institutional Review Board of our institution. Peripheral blood mononuclear cells were isolated by density gradient centrifugation, followed by culturing in EGM-2MV medium. Nonadherent cells were removed 3 days after cell culture and fresh medium was changed every 2 days. Adherent EPCs were identified by their formation of a cobblestonelike morphology and specific expression of surface markers (CD34, CD31, VEGF recepter (R)-2, and von Willebrand factor [vWF]) using flow cytometry. EPCs from passages three to four were harvested for the following experiments. To alter the expression of miR-22-3p, we applied miR-22-3p inhibitor to EPCs with lipofectamine 2000 (Invitrogen; Carlsbad, CA, USA) and tested the transfection effect by quantitative real-time polymerase chain reaction (qRT-PCR).
CELL PROLIFERATION ASSAY:
The proliferative capacity of EPCs was evaluated by Cell-Counting Kit 8 (CCK-8) assay. A total of 2×104 cells was seeded into each well of a 24-well plate and cultured in an incubator for 72 h. After that, 10 μL of CCK-8 solution (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added to each well and incubated for 1 h. The absorbance value of each well was detected at a wavelength of 450 nm at 24, 48, and 72 h after incubation. All experiments were conducted in triplicate.
TRANSWELL ASSAY:
For the transwell assay, 1×105 cells were suspended in basic medium (EBM-2MV) and seeded into the upper chamber of a 24-well transwell plate. The EBM-2MV medium with 20% fetal bovine serum was added into the lower chamber. After 24 h, the cells migrated into the lower chamber were stained with 0.5% crystal violet and the number of migrated cells was counted under a light microscope. All experiments were performed in triplicate.
SCRATCH WOUND ASSAY:
First, 1×106 cells were plated onto a 6-well plate and grown to 80% confluence. The monolayer was scratched with a 20-μL pipette tip and washed with phosphate-buffered saline to remove the detached cells. Then the adherent cells were incubated with EBM-2MV medium in a 5% CO2 cell culture incubator. The plates were photographed at 0 and 48 h after scratching, respectively. Each experiment was done in triplicate.
QRT-PCR ANALYSIS:
Total RNA was isolated using TRIzol reagent (Invitrogen) and reverse-transcribed using Moloney murine leukemia virus reverse transcription kit (Promega, Madison, WI, USA). qRT-PCR was conducted by using SYBR® Green qPCR supermixes (Bio-Rad, Berkeley, CA, USA). Mature miRNA expression analysis was performed with miRNA real-time PCR quantitation kit (Applied Biosystems, Foster City, CA, USA). Glyceraldehyde 3-phosphate (GAPDH) and U6 served as reference controls. PCR primers were as follows:
WESTERN BLOT ANALYSIS:
EPCs were collected with radioimmunoprecipitation assay lysis buffer and phenylmethylsulfonyl fluoride, quantified by a bicinchoninic acid protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Then the mixed buffer was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride (PVDF) membranes, which were subsequently blocked in 10% nonfat milk-Tris-buffered saline with Tween 20 (TBST) for 1 h and incubated in primary antibody overnight. After that, the PVDF membranes were incubated with horseradish peroxidase-conjugated secondary antibody. Quantification of band intensities was analyzed with Image J software.
LUCIFERASE REPORT ASSAY:
The sequence of 3′-untranslated region (UTR) of OC1 with putative binding sites was cloned and inserted into the
SERUM RNA EXTRACTION:
Peripheral blood samples were collected in ethylenediaminetetraacetic acid-anticoagulant tubes and centrifuged at 3000 revolutions per min for 15 min at 4°C. Small RNAs were extracted from 500 μL of plasma supernatant using a miR-PARIS kit according to the manufacturer’s instructions.
TREATMENT AND FOLLOW-UP:
All the patients recruited in this study were treated with standard anticoagulation therapy in combination with thrombolysis treatment. Peripheral blood sample were collected and the venous patency monitored on each follow-up by duplex sonography. The expression level of miR-22-3p was tested and compared between patients with and without recurrent DVT.
STATISTICAL ANALYSIS:
Continuous variables are presented as mean and standard deviation and discrete outcomes are presented as percentages. Student’s
Results
MIR-22-3P IS UPREGULATED IN EPCS DERIVED FROM DVT:
To identify the harvest cells, EPCs were confirmed by their morphology and the expression of specific cell markers including CD34, CD31, VEGFR-2, and vWF (Supplementary Figure 1). Then we tested miR-22-3p expression level in EPCs both from healthy volunteers and DVT patients. Gene expression analysis showed that miR-22-3p was upregulated in EPCs from DVT patients compared with cells from healthy volunteers (Figure 1).
SUPPRESSION OF MIR-22-3P LEADS TO PROLIFERATION AND MIGRATION OF EPCS:
To verify the effect of miR-22-3p on proliferation and migration of EPCs, the expression level of miR-22-3p was altered by transfecting with miR-22-3p inhibitor or its negative control. Next, transfected cells were examined using a series of in vitro assays. Results from CCK-8 assay showed that inhibition of miR-22-3p increased the proliferative capability of EPCs as compared with the control group (Figure 2A). Furthermore, we verified that reduced expression of miR-22-3p also promoted migration capacity with transwell assay and scratch wound assay (Figure 2B–2E). Taken together, these data suggest the regulatory function of miR-22-3p on proliferation and migration of EPCs.
OC1 IS REGULATED BY MIR-22-3P:
It is known that miRNAs participate in RNA splicing and posttranscriptional regulation via binding 3′-UTR of target messenger (m)RNA. To figure out the potential mechanism of miR-22-3p in regulating EPCs, bioinformatic analysis was used for searching for the target gene. Thus, we searched multiple databases such as miRDB, Miranda, and TargetScan and our results revealed that OC1 was a candidate target gene for miR-22-3p (Figure 3A). Next, we examined the direct interaction between miR-22-3p and OC1 by using luciferase report assay. Analysis showed a decrease of luciferase activity in the presence of miR-22-3p, indicating binding of miR-22-3p to OC1 3′-UTR (Figure 3B). Consistently, protein expression level of OC1 was increased after inhibition of miR-22-3p (Figure 3C), confirming the bioinformatics prediction.
MIR-22-3P NEGATIVELY REGULATES THE EXPRESSION OF VEGFA IN EPCS:
Previous studies have suggested that VEGFA played an important role in regulation of endothelial cells. In addition, OC1 was believed to be involved in VEGFA transcriptional expression. Here, we hypothesized the contribution of VEGFA in regulating EPC biological function. Gene analysis revealed that inhibition of miR-22-3p induced upregulation of VEGFA (Figure 4A). Moreover, western blot analysis also confirmed an increased expression of VEGFA at the protein level (Figure 4B, 4C). We also applied OC1 small interfering RNA into EPCs and found the opposite effect (Figure 4D–4F). Taken together, these results demonstrated that miR-22-3p regulated EPC function via targeting VEGFA.
MIR-22-3P IS ASSOCIATED WITH PROGNOSIS IN VT:
It was found that miRNAs could function as biomarkers for prognosis of diseases. We therefore investigated the correlation of miR-22-3p and prognosis of VT. Serum miR-22-3p was collected from patients during follow-up. The venous patency was detected by duplex sonography to monitor reoccurrence of DVT. A total of 58 patients was included in this study from January 2013 to August 2018. The baseline characteristics of patients are listed in Table 1. Eighteen cases were diagnosed with DVT recurrence during follow-up (Figure 5A, 5B). We also compared the expression of miR-22-3p between patients with and without recurrent DVT. Analysis showed a higher expression level of miR-22-3p in recurrent DVT compared with those without reoccurrence (Figure 5C).
Discussion
In this study, we found that miR-22-3p contributed to the dysfunction of EPCs in the setting of VT and the inhibition of miR-22-3p led to significant increase of OC1 and VEGFA expression at the protein level. We also showed a new form of communication between miR-22-3p and OC1. Furthermore, the aberrant overexpression of miR-22-3p indicated a higher DVT recurrence risk for patients.
DVT occurs when a blood clot forms in one or more of the deep veins in the body, and can cause leg pain or swelling. Epidemiological studies have demonstrated that over 10 million DVT cases occur annually worldwide [11]. The standard treatment regimens include systematic anticoagulation, thrombolysis, and physical therapy. However, standard treatment has limited effect on repairing the injured intima and restoring the function of venous valves, leading to frequent complication of postthrombotic syndrome (PTS). It is estimated that 20–50% of patients develop PTS within 2 years of onset [12]. Apart from PTS, recurrence of DVT is another potential consequence. Previous research showed that around 30% of patients experience a recurrence within 10 years after an initial DVT [13,14]. EPCs have been shown to participate in blood vessel formation, vascular intima repair, and re-endothelialization of denuded vessels [15]. Furthermore, in many instances, circulating EPC concentrations have been enumerated and correlated to a disease in an effort to serve as a biomarker for disease detection or staging [16,17]. Previous studies have revealed that EPCs would migrate into a thrombus and participate in the recanalization [7,18]. However, some researchers found that EPCs were dysfunctional in DVT patients [19]. It is believed that the pathological circumstance of VT impairs normal biological function of EPCs. Thus, in the present study, we focused on the mechanism behind the phenomenon.
MiRNAs are short noncoding nucleotides that can regulate the expression of a large number of genes and have been therefore implicated in numerous physiological and pathological processes, including migration, angiogenesis, and proliferation. Recent papers have elucidated a critical role of miRNA in EPCs. Some specific miRNAs have been correlated with a change in the posttranscriptional editing of target mRNA that results in subsequent effects on biological function. Kong et al. [18] revealed that miR-483-3p contributed to EPC dysfunction via serum response factor. In addition, downregulation of let-7e-5p is associated with dysfunction of EPCs via targeting of the Fas ligand gene [7]. The results published by Wang et al. showed that miR-150 promoted angiogenesis and proliferation of EPCs by targeting SRC kinase signaling inhibitor 1 [20]. In line with previous studies, we also found the miRNA-induced mechanism in regulating EPCs. One of the most important observations in this research is the confirmation of a novel miR-22-3p-dependent regulation of EPC function. First, we found the aberrant expression of miR-22-3p in EPCs from DVT patients. Meanwhile, inhibition of miR-22-3p increased migration and proliferation of EPCs via its target gene OC1. Interestingly, the suppression of miR-22-3p also induced the upregulation of VEGFA, which is believed to be the downstream target of OC1. In addition, previous studies have confirmed the role of specific miRNAs in screening and predicting prognosis of diseases. Xue et al. [21] reported that circulating miR-17-5p, miR-126-5p, and miR-145-3p were novel biomarkers for diagnosis of acute myocardial infarction. Here, we also find that higher expression of miR-22-3p is associated with recurrent DVT during follow-up, which is beneficial for clinical prognosis.
Conclusions
In summary, we demonstrated the role of miR-22-3p in regulating the biological function of EPCs via targeting OC1 and therefore affecting the expression of VEGFA. Furthermore, serum miR-22-3p had unfavorable trends of recurrent risk and could be monitored for prognosis of DVT. Our study identifies a mechanism-based biological repair strategy for DVT and provides clinically relevant data on prognosis after VT.
Figures
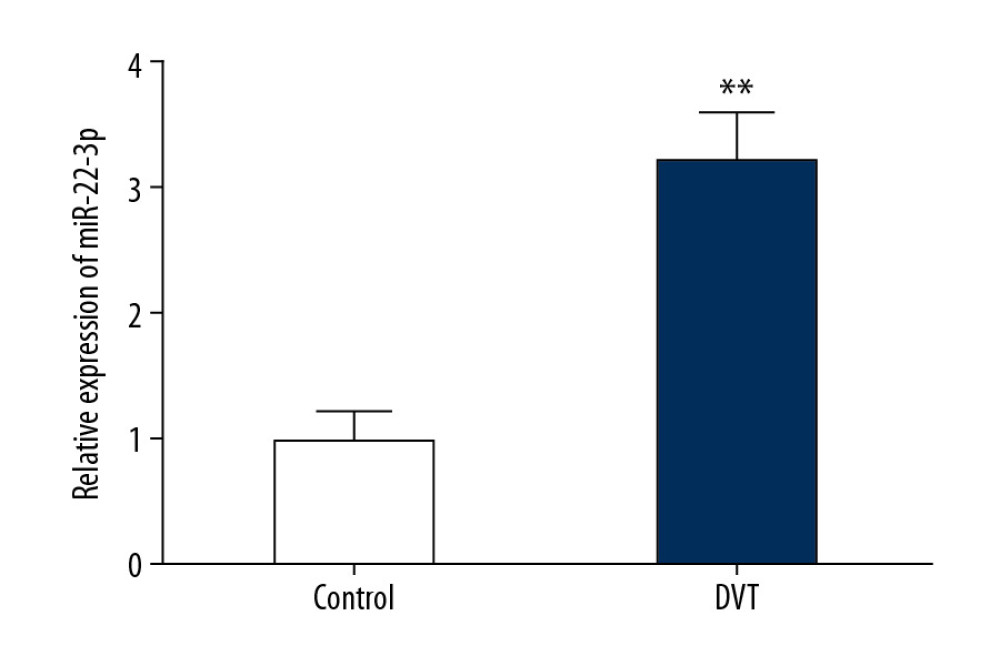 Figure 1. Expression of miR-22-3p in endothelial progenitor cells (EPCs) from deep vein thrombosis (DVT) patients and healthy volunteers. (** P<0.01).
Figure 1. Expression of miR-22-3p in endothelial progenitor cells (EPCs) from deep vein thrombosis (DVT) patients and healthy volunteers. (** P<0.01). 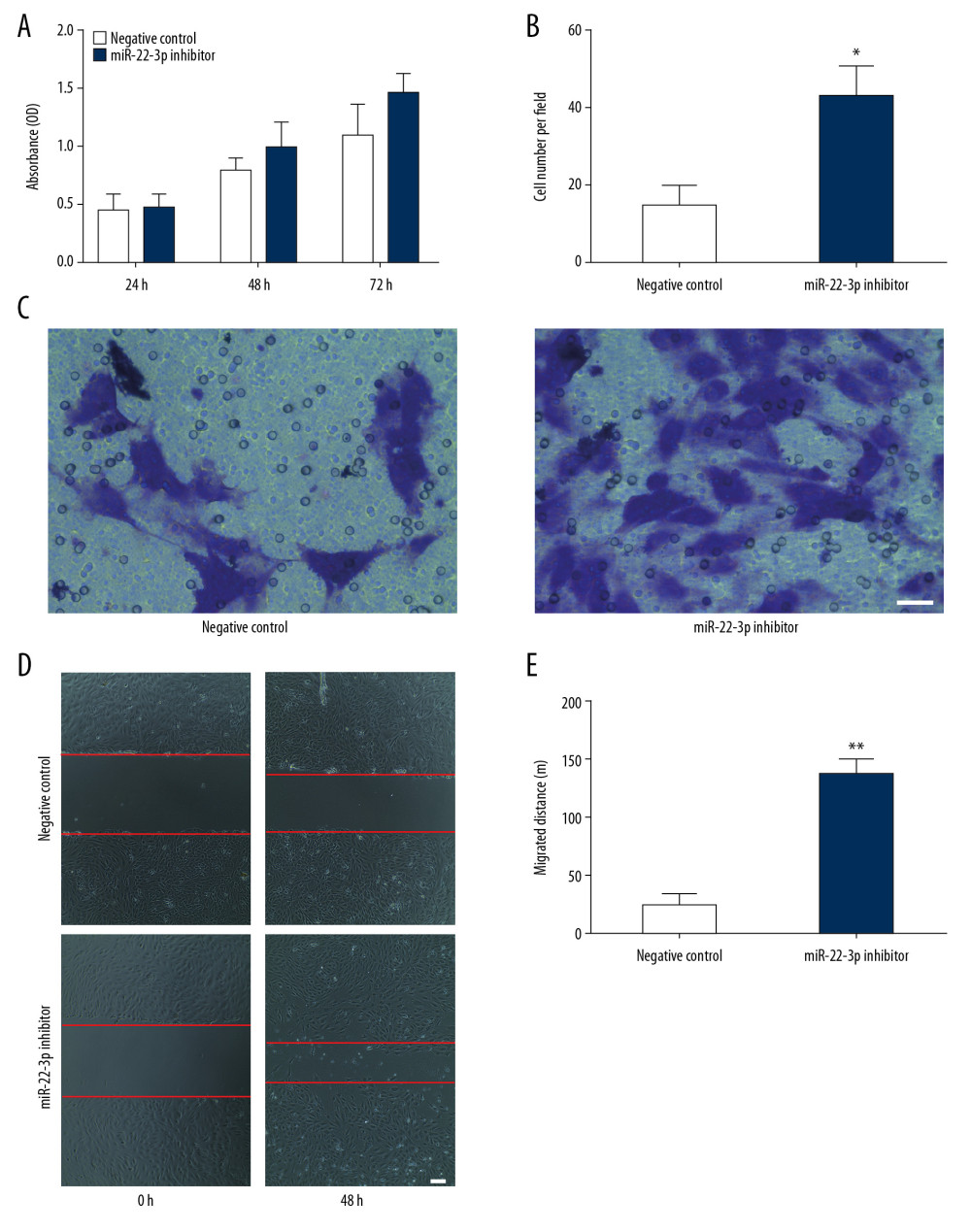 Figure 2. Effect of miR-22-3p on the biological function of endothelial progenitor cells (EPCs). (A) Proliferation assay showing knockdown of miR-22-3p promoted cell proliferation. (B) Statistical analysis of transwell assay revealed that inhibition of miR-22-3p increased migration of EPCs. (C) Representative microscopic images of cells that invaded through the transwell. (D) EPCs transfected with miR-22-3p inhibitor showed more migrated distance. (E) Statistical results from wound-healing assay. * P<0.05; ** P<0.01.
Figure 2. Effect of miR-22-3p on the biological function of endothelial progenitor cells (EPCs). (A) Proliferation assay showing knockdown of miR-22-3p promoted cell proliferation. (B) Statistical analysis of transwell assay revealed that inhibition of miR-22-3p increased migration of EPCs. (C) Representative microscopic images of cells that invaded through the transwell. (D) EPCs transfected with miR-22-3p inhibitor showed more migrated distance. (E) Statistical results from wound-healing assay. * P<0.05; ** P<0.01. 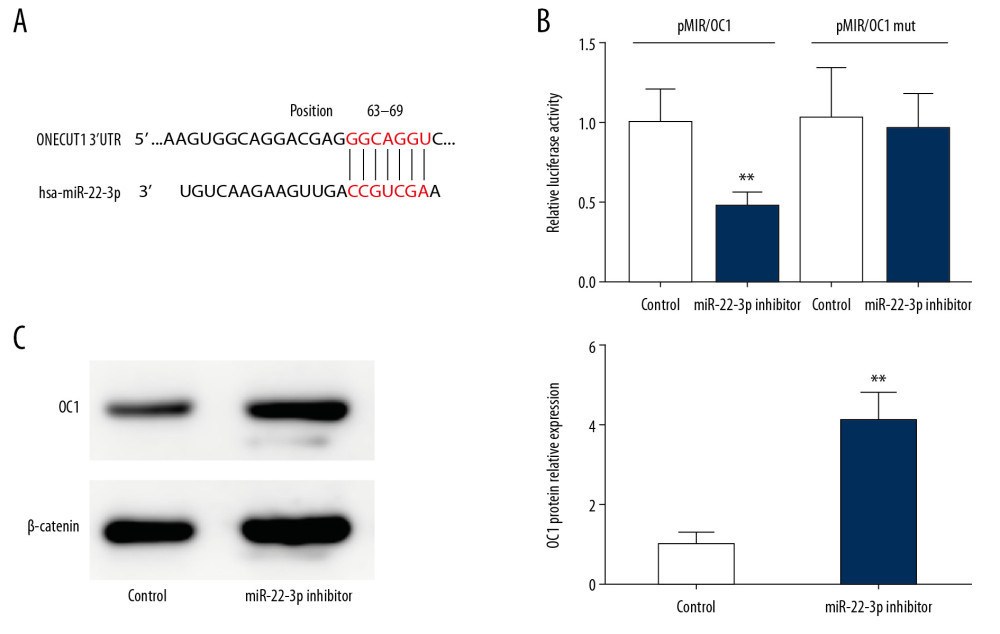 Figure 3. Onecut 1 (OC1) is negatively regulated by miR-22-3p. (A) Bioinformatic analysis predicted the 3′-untranslated region (UTR) of OC1 with the potential binding sites of miR-22-3p. (B) Luciferase report assay showed the activity in different groups. (C) Western blot analysis revealed the protein expression of OC1. ** P<0.01.
Figure 3. Onecut 1 (OC1) is negatively regulated by miR-22-3p. (A) Bioinformatic analysis predicted the 3′-untranslated region (UTR) of OC1 with the potential binding sites of miR-22-3p. (B) Luciferase report assay showed the activity in different groups. (C) Western blot analysis revealed the protein expression of OC1. ** P<0.01. 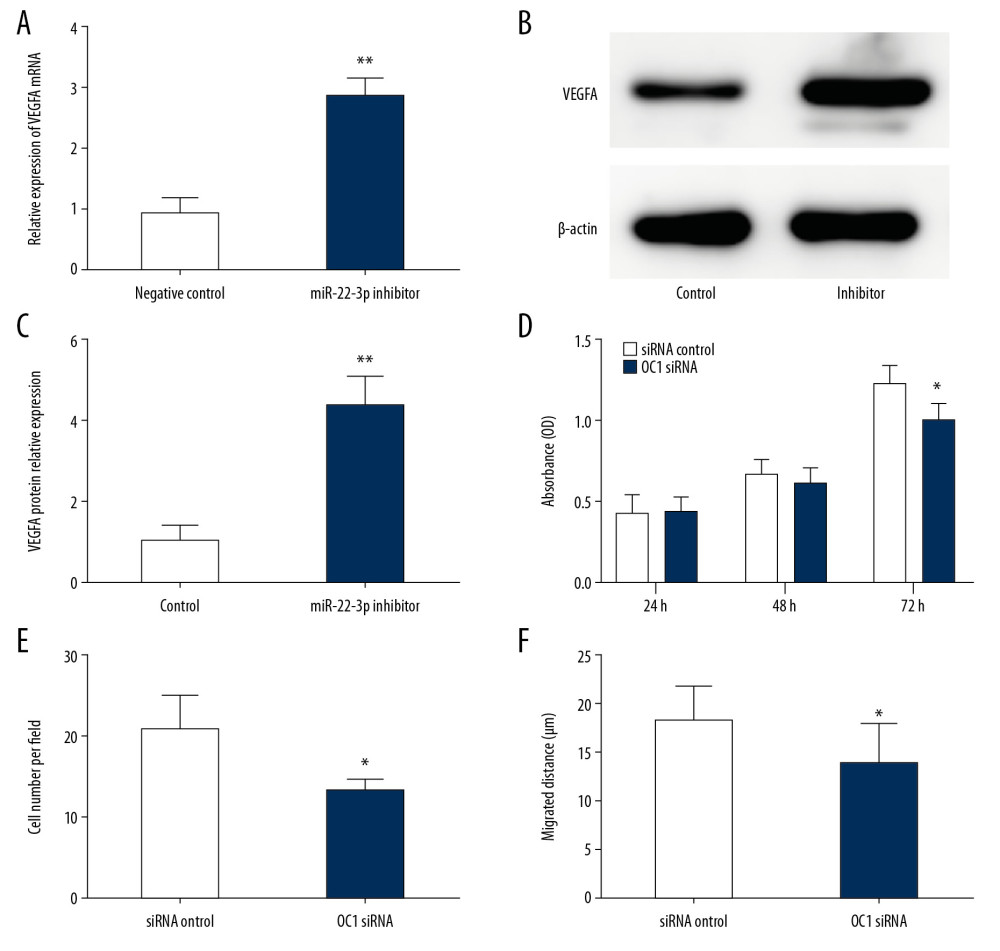 Figure 4. MiR-22-3p negatively regulates the expression of vascular endothelial growth factor A (VEGFA). (A) VEGFA messenger ribonucleic acid (mRNA) expression detected by quantitative real-time polymerase chain reaction (qRT-PCR) in different groups. (B) VEGFA protein expression analyzed by Western blot in different groups. (C) Quantitative analysis of VEGFA protein level in endothelial progenitor cells (EPCs) transfected with different duplexes. (D) Cell-Counting Kit 8 (CCK-8) assay showed the proliferative ability of EPCs in different groups. (E) Transwell assay revealed migration of EPCs transfected with either small interfering (si)RNA control or OC1 siRNA. (F) Wound-healing assay detected the migration ability of EPCs in different groups. * P<0.05; ** P<0.01.
Figure 4. MiR-22-3p negatively regulates the expression of vascular endothelial growth factor A (VEGFA). (A) VEGFA messenger ribonucleic acid (mRNA) expression detected by quantitative real-time polymerase chain reaction (qRT-PCR) in different groups. (B) VEGFA protein expression analyzed by Western blot in different groups. (C) Quantitative analysis of VEGFA protein level in endothelial progenitor cells (EPCs) transfected with different duplexes. (D) Cell-Counting Kit 8 (CCK-8) assay showed the proliferative ability of EPCs in different groups. (E) Transwell assay revealed migration of EPCs transfected with either small interfering (si)RNA control or OC1 siRNA. (F) Wound-healing assay detected the migration ability of EPCs in different groups. * P<0.05; ** P<0.01. 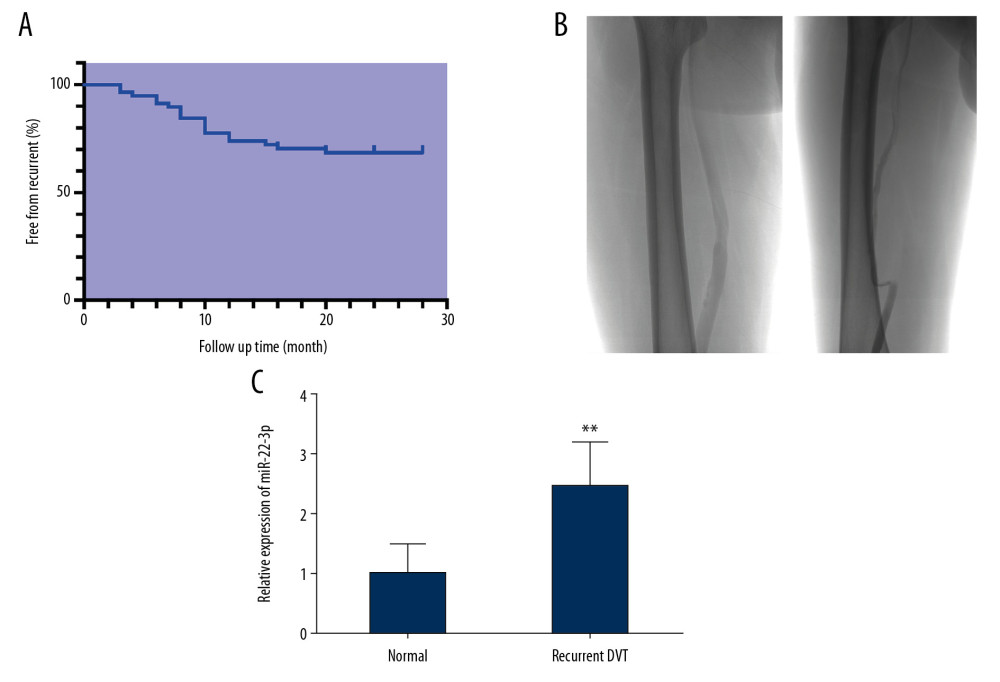 Figure 5. MiR-22-3p is associated with prognosis of deep vein thrombosis (DVT). (A) Kaplan-Meier curve revealed recurrence-free survival. (B) Representative digital subtraction angiography (DSA) images for patent vein (left) and recurrent venous thrombosis (VT, right). (C) Comparison of miR-22-3p expression in both normal and recurrent deep (D)VT patients. ** P<0.01.
Figure 5. MiR-22-3p is associated with prognosis of deep vein thrombosis (DVT). (A) Kaplan-Meier curve revealed recurrence-free survival. (B) Representative digital subtraction angiography (DSA) images for patent vein (left) and recurrent venous thrombosis (VT, right). (C) Comparison of miR-22-3p expression in both normal and recurrent deep (D)VT patients. ** P<0.01. 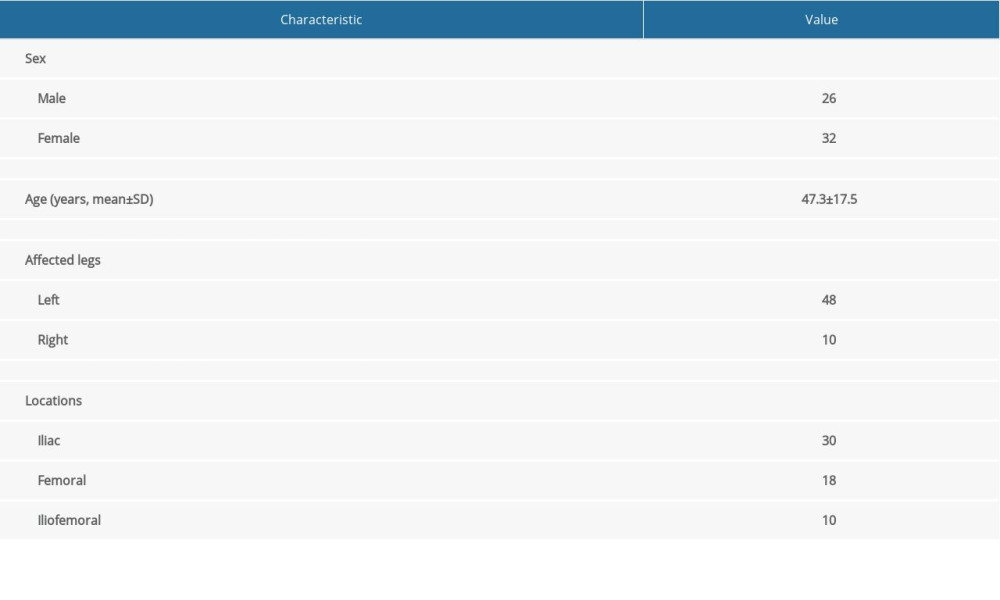
References
1. Chong MSK, Ng WK, Chan JKY, Concise review: Endothelial progenitor cells in regenerative medicine: Applications and challenges: Stem Cells Transl Med, 2016; 5(4); 530-38
2. Werner N, Kosiol S, Schiegl T, Circulating endothelial progenitor cells and cardiovascular outcomes: N Engl J Med, 2005; 353(10); 999-1007
3. Ambros VR, The functions of animal microRNAs: Nature, 2004; 431(7006); 350-55
4. Lin S, Gregory RI, MicroRNA biogenesis pathways in cancer: Nat Rev Cancer, 2015; 15(6); 321-33
5. Lu Y, Thavarajah T, Gu W, Cai J, Xu Q, Impact of miRNA in atherosclerosis: Arterioscler Thromb Vasc Biol, 2018; 38(9); e159-70
6. Goerke SM, Kiefer LS, Stark GB, miR-126 modulates angiogenic growth parameters of peripheral blood endothelial progenitor cells: Biol Chem, 2015; 396(3); 245-52
7. Kong L, Du X, Hu N, Downregulation of let-7e-5p contributes to endothelial progenitor cell dysfunction in deep vein thrombosis via targeting FASLG: Thromb Res, 2016; 138(138); 30-36
8. Li W, Zhou D, Sun L, LncRNA WTAPP1 promotes migration and angiogenesis of endothelial progenitor cells via MMP1 through microRNA 3120 and Akt/PI3K/autophagy pathways: Stem Cells, 2018; 36(12); 1863-74
9. Huang S, Wang M, Wu W, Mir-22-3p inhibits arterial smooth muscle cell proliferation and migration and neointimal hyperplasia by targeting HMGB1 in arteriosclerosis obliterans: Cellular Physiol Biochem, 2017; 42(6); 2492-506
10. Ma J, Shang S, Wang J, Identification of miR-22-3p, miR-92a-3p, and miR-137 in peripheral blood as biomarker for schizophrenia: Psychiatry Res, 2018; 265; 70-76
11. Raskob GE, Angchaisuksiri P, Blanco AN, Thrombosis: A major contributor to global disease burden: Arterioscler Thromb Vasc Biol, 2014; 34(11); 2363-71
12. Mazzolai L, Aboyans V, Ageno W, Diagnosis and management of acute deep vein thrombosis: A joint consensus document from the European Society of Cardiology working groups of aorta and peripheral vascular diseases and pulmonary circulation and right ventricular function: Eur Heart J, 2018; 39(47); 4208-18
13. Beckman MG, Hooper WC, Critchley SE, Ortel TL, Venous thromboembolism: A public health concern: Am J Prev Med, 2010; 38(4); S495-501
14. White RH, The epidemiology of venous thromboembolism: Circulation, 2003; 107(23 Suppl 1); I4-8
15. Zilla P, Bezuidenhout D, Human P, Prosthetic vascular grafts: Wrong models, wrong questions and no healing: Biomaterials, 2007; 28(34); 5009-27
16. Bakogiannis C, Tousoulis D, Androulakis E, Circulating endothelial progenitor cells as biomarkers for prediction of cardiovascular outcomes: Curr Med Chem, 2012; 19(16); 2597-604
17. Sen S, McDonald SP, Coates PT, Bonder CS, Endothelial progenitor cells: Novel biomarker and promising cell therapy for cardiovascular disease: Clin Sci (Lond), 2011; 120(7); 263-83
18. Kong L, Hu N, Du X, Upregulation of miR-483-3p contributes to endothelial progenitor cells dysfunction in deep vein thrombosis patients via SRF: J Transl Med, 2016; 14; 23
19. Alvaradomoreno JA, Hernandezlopez R, Chavezgonzalez A, Endothelial colony-forming cells: Biological and functional abnormalities in patients with recurrent, unprovoked venous thromboembolic disease: Thromb Res, 2016; 137; 157-68
20. Wang W, Zhu X, Du X, MiR-150 promotes angiogensis and proliferation of endothelial progenitor cells in deep venous thrombosis by targeting SRCIN1: Microvasc Res, 2019; 123; 35-41
21. Xue S, Liu D, Zhu W, Circulating miR-17-5p, miR-126-5p and miR-145-3p are novel biomarkers for diagnosis of acute myocardial infarction: Front Physiol, 2019; 10; 123
Figures
Figure 1. Expression of miR-22-3p in endothelial progenitor cells (EPCs) from deep vein thrombosis (DVT) patients and healthy volunteers. (** P<0.01).Figure 2. Effect of miR-22-3p on the biological function of endothelial progenitor cells (EPCs). (A) Proliferation assay showing knockdown of miR-22-3p promoted cell proliferation. (B) Statistical analysis of transwell assay revealed that inhibition of miR-22-3p increased migration of EPCs. (C) Representative microscopic images of cells that invaded through the transwell. (D) EPCs transfected with miR-22-3p inhibitor showed more migrated distance. (E) Statistical results from wound-healing assay. * P<0.05; ** P<0.01.Figure 3. Onecut 1 (OC1) is negatively regulated by miR-22-3p. (A) Bioinformatic analysis predicted the 3′-untranslated region (UTR) of OC1 with the potential binding sites of miR-22-3p. (B) Luciferase report assay showed the activity in different groups. (C) Western blot analysis revealed the protein expression of OC1. ** P<0.01.Figure 4. MiR-22-3p negatively regulates the expression of vascular endothelial growth factor A (VEGFA). (A) VEGFA messenger ribonucleic acid (mRNA) expression detected by quantitative real-time polymerase chain reaction (qRT-PCR) in different groups. (B) VEGFA protein expression analyzed by Western blot in different groups. (C) Quantitative analysis of VEGFA protein level in endothelial progenitor cells (EPCs) transfected with different duplexes. (D) Cell-Counting Kit 8 (CCK-8) assay showed the proliferative ability of EPCs in different groups. (E) Transwell assay revealed migration of EPCs transfected with either small interfering (si)RNA control or OC1 siRNA. (F) Wound-healing assay detected the migration ability of EPCs in different groups. * P<0.05; ** P<0.01.Figure 5. MiR-22-3p is associated with prognosis of deep vein thrombosis (DVT). (A) Kaplan-Meier curve revealed recurrence-free survival. (B) Representative digital subtraction angiography (DSA) images for patent vein (left) and recurrent venous thrombosis (VT, right). (C) Comparison of miR-22-3p expression in both normal and recurrent deep (D)VT patients. ** P<0.01. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387