22 June 2021: Clinical Research
Cortical and Subcortical Brain Volume Alterations Following Endurance Running at 38.6 km and 119.2 km in Male Athletes
Rabia Hurrem Ozdurak Singin1ABCDEFG, Serkan Duz2ABCDEFG*, Murat Kiraz3AACCDDEEDOI: 10.12659/MSM.926060
Med Sci Monit 2021; 27:e926060






Abstract
BACKGROUND: Although several studies have shown that ultramarathon running causes severe physical and mental stress and harms organ systems, its effect on brain tissue remains unclear. The purpose of this study was to investigate the volumetric change of cortical and subcortical brain structures following 38.6-km and 119.8-km mountain races.
MATERIAL AND METHODS: A total of 23 healthy male runners (age, 49.05±5.99 years) were classified as short-trail (ST; n=9) and ultra-trail (UT; n=14) endurance running. Pre- and post-test scanning of brain tissue was performed by using a 3-Tesla magnetic resonance imaging (MRI). Pre- and post-race differences in cortical and subcortical volumes in the ST and UT groups were separately determined by Wilcoxon signed-rank test.
RESULTS: Cortical gray matter (GM) and cerebral GM volume significantly increased after the race in both ST and UT groups, whereas the volume of the thalamus, caudate, pallidus, and hippocampus significantly increased only in the UT group. Cerebrospinal fluid (CSF) and white-matter (WM) volumes did not change after endurance running and remained unaltered in both groups.
CONCLUSIONS: Endurance running has a site-specific acute effect on cortical and subcortical structures and may attenuate GM volume decrease in older adult male athletes. The increased volume of subcortical structures might be a response of physical exercise and additional physical stress experienced by ultramarathon runners.
Keywords: Diffusion Magnetic Resonance Imaging, Neuroimaging, Running, athletes, Gray matter, Magnetic Resonance Imaging, Marathon Running, Organ Size, Physical Endurance, Reference Values, white matter
Background
The beneficial effects of regular physical exercise on human health and quality of life have been widely accepted and documented, with relevant evidence that it significantly improves inactivity-related diseases such as hypertension, obesity, dyslipidemia, and cardiovascular disease [1–3]. Studies have proven that strenuous exercise performed over a certain intensity or workload harms the health and is associated with serious damage in many organs and systems [4,5]. Short-term endurance exercise has been reported to have severe adverse effects on the cardiorespiratory system [6], hormonal status [7], immune function [8], electrolyte metabolism [9], and muscle damage [10].
Running is one of the world’s most popular sports performed for health, fitness benefits, enjoyment, and socialization [11]. Therefore, it is performed by millions of enthusiasts at various distances, even despite their advancing age [12,13]. Amateur runners start their running experience with short distances that may gradually increase up to ultramarathons, which are races longer than the traditional 42.2 km marathon. Ultramarathon races are performed at various distances, ranging from 50 km [14] up to continental races, but the most preferred distance seems to be over 100 km [15–17].
Most recreational ultramarathon runners reach their peak performance in their 40s and are willing to push their limits in extreme conditions that make them prone to diverse medical complications that vary from mild blisters and stress fractures to severe cardiac arrest or stroke [18–21]. Cardiac arrest [22], hyponatremia [23,24], increased arterial stiffness [25], and kidney dysfunction [26] were reported after ultramarathons, but the effect of physical overexertion on the brain remains unclear.
Recent studies clearly show the beneficial effects of moderate exercise on brain tissue, demonstrating that it can prevent and even reverse age-dependent decline in brain tissue and thereby improve cognitive functions of elderly people with dementia [27–29]. Thus, brain atrophy in the elderly can be prevented or slowed by aerobic exercise as a result of enhanced synaptic plasticity in the hippocampus and increased cell poliferation [30]. Moreover, increased blood flow to the hippocampus in adults is known to improve memory functions and increase neurogenesis [31]. Wood et al [32] reported increased cortical GM volume as a long-term adaptation to ultramarathon running in athletes with running experience of over 30 years.
To date, few studies have focused on the effects of ultramarathon running on the brain, and only 2 studies have focused on acute effects on cerebral volumetric changes during endurance running. The first prospective MRI study reported GM volume decline after a 4487-km multistage continental ultramarathon race [33]. On the other hand, Herm et al [34] found that WM volume was not altered after marathon running when compared to controls, and concluded that endurance running did not cause brain injury. However, results of these MRI studies are not comparable due to use of different measurement techniques and the nature and distance of the ultramarathon. Thus, more detailed research is needed to understand the effect and mechanism of ultramarathon running on brain tissue.
The purpose of this study was to identify the acute effects of marathon and ultramarathon running on cortical and subcortical structures in male runners by using a high-resolution 3-Tesla MRI.
Material and Methods
PARTICIPANTS:
All runners signed an informed consent form before participation. The study adhered to the Helsinki Declaration, and Clinical Research Ethics Committee approval was obtained from the local university to eliminate ethical concerns (approval no. 83/09-10-2019). The quasi-experimental study was performed with nonrandomized groups and a pre- and post-test design.
Runners who enrolled to participate in the Salomon Cappadocia Ultra-trail 2019 event were contacted a few weeks before the race and requested to participate in the research. Enrollment criteria included either having a valid medical report stating that there was no health hazard for the person to participate in an ultramarathon run or having a permit approved by any sports federation. A total of 27 healthy amateur male runners (age, 49.05±5.99 years) were eligible for inclusion (age, handedness, health status), and were non-randomly grouped as short-trail (ST; n=12) or ultra-trail (UT; n=15) according to their registration preference for the race. We excluded participants who consumed alcohol or tobacco, had any kind of neurologic, psychologic, or metabolic diseases, or who had a history of traumatic brain injury.
DESIGN:
The measurements were performed 1 day before the race and immediately after the race. The Salomon Cappadocia Ultra-trail race is monitored by the International Trail Race Association (ITRA), and guarantees certain standards. Athletes were allowed to consume food, water, and hypotonic drinks at check points that had to be reached within certain time limits. ST and UT routes are given in detail in Figures 1 and 2, respectively.
The total altitude changes for the 38.6 km ST and 119.8 km UT races were 1120 m and 3730 m, respectively. Average temperature (day=16.5±3.1°C; night=8.8±2.1°C; 6–21°C), humidity (50±1.41%), and wind speed (13 km/h) were checked from AccuWeather (AccuWeather Superior Accuracy™), the Wet Bulb Globe Temperature index was calculated as 12.2°C, and the heat index was 17.23°C.
DEMOGRAPHIC AND ANTHROPOMETRIC DATA: Demographic data were obtained via questionnaire during pre-test assessment, including personal information, medical conditions, and training history. Height, body weight, and body-mass index (BMI) of the athletes were assessed according to the standardized anthropometric procedures [35].
MRI ACQUISITION:
Two brain scans were performed for each athlete by the same radiologist, 1 day before and immediately after the race. The time of the pre-race MRI scan of each athlete was planned according to the estimated time of arrival, which was calculated based on the athlete’s prior race performance. Brain scans were obtained using a Siemens 3.0 Tesla MRI (Magnetom Skyra, Siemens Healthcare, Erlangen, Germany) at the National Research and Education City Hospital. The 3D sagittal magnetization-prepared rapid-acquisition gradient-echo (MPRAGE) sequence (sagittal, repetition time (TR)=2300 ms, echo time (TE)=3.4 ms, field of view (FOV)=250 mm2, matrix: 256×256, flip angle 7°, slice thickness=1 mm) of T1-weighted images was obtained in supine position.
MRI PROCESSING: VolBrain (http://volbrain.upv.es) cloud-computing software was used for processing T1-weighted MR images. Cerebrospinal fluid (CSF), brainstem, gray-matter (GM), white-matter (WM), and total brain volumes of cortical and GM of subcortical structures, including thalamus, caudate, globus pallidus, hippocampus, accumbens, putamen, and amygdala were demonstrated as mean±SD (min–max). Prerequisite steps of the pipeline are explained in detail elsewhere [36].
STATISTICAL ANALYSIS:
Whether the data showed normal distribution was tested by using the Shapiro-Wilk test by using the IBM statistical package program (SPSS version 25.0, IBM Corp.: Armonk, NY, USA). The Wilcoxon signed-rank test was used to determine differences in brain volume after the race in ST and UT groups, separately. The alpha value was accepted as p<0.05.
Results
DESCRIPTIVE PARTICIPANT STATISTICS:
A total of 1207 runners (715 male and 492 female) departed for the ST (Figure 1), and 363 runners (324 male and 39 female) departed for the UT (Figure 2) route. Twelve runners in the ST and 15 runners in the UT were scanned before the race. Only 9 runners in ST and 14 runners in UT were scanned after the race. Four runners were excluded because of muscle injuries (n=2) or drop-out (n=1), and 1 runner in the UT group refused to participate in post-race measurements.
All participants were experienced but non-professional athletes with an average running history at least 2 years and were identical in terms of weight, height and BMI in both ST and UT groups. Although average running distance per year and per week was greater in the UT group compared to the ST group, both groups were adapted long-term to endurance running. Although the average training volume was twice as high in the UT group compared to the ST group, performance expressed as running speed in training and race did not differ between the ST and UT groups. Descriptive data and characteristics of athletes are given in Table 1.
VOLUMETRIC CHANGES IN CORTICAL AND SUBCORTICAL BRAIN REGIONS FOLLOWING THE MARATHON AND THE ULTRAMARATHON RACE:
Pre- and post-race brain volumes of cortical and subcortical structures in ST (Table 2) and UT (Table 3) groups were evaluated, separately. According to the Wilcoxon signed-rank test, total brain volume increased significantly after the race in the UT group (Z=−2.23; p=0.026), but not in the ST group (Z=−1.36; p=0.173) when pre-race values were compared with post-race values (Figure 3).
A statistically significant increase in tissue volume was observed in total GM volume in both the ST (Z=−2.31; p=0.021) and the UT (Z=−2.86; p=0.004) groups (Figure 4), whereas the volume of WM, CSF and brainstem did not change between pre- and post-race values in either group (Tables 2, 3).
The cerebral GM volume was significantly higher after the race in both ST (Z=−2.31; p=0.021) and UT (Z=−2.86; p=0.004) groups (Figure 5).
On the other hand, total cerebral volume increased significantly between pre- and post-race in the UT group (Z=−2.04; p=0.041) but not in the ST group (Z=−1.72; p=0.086) (Figure 6).
Volume of cerebrum WM and cerebellum remained same and did not change in either group (Tables 2, 3).
Volumes of the thalamus (Z=−2.61; p=0.009) (Figure 7), caudate (Z=−2.41; p=0.041) (Figure 8), pallidus (Z=−2.64; p=0.008) (Figure 9), and hippocampus (Z=−3.31; p=0.001) (Figure 10) increased significantly after the UT race, whereas putamen, amygdala, and accumbens volumes did not change in the UT group. In the ST group, volumes of all subcortical structures remained unchanged when pre-race values were compared to post-race values (Table 2).
Discussion
This study assessed the acute impact of marathon and ultramarathon running on brain volume by using high-resolution 3-Tesla MRI scanning in older adult male athletes. We assumed that running distance may play a differential role and cause various physical and mental stress, giving rise to changes in the brain structures of male runners. We found a clear increase in total cerebral GM volume after the marathon and ultramarathon, whereas CSF, brainstem, and WM volumes did not change in either group. Moreover, volumetric increases in the thalamus, caudate, pallidus, and hippocampus after ultramarathon, but not marathon, running suggests that the response of brain tissue volume depends on running distance and exercise amount.
The acute increase in the GM, but not WM, volume after endurance running demonstrates that GM is more sensitive to exercise compared to WM tissue. This finding is similar to the results of the study focusing on long-term adaptation of ultramarathon in brain tissue of experienced athletes. Runners with experience of over 30 years had higher total GM volumes compared to physically active athletes, but WM volume did not differ between groups [32]. There is evidence that brain morphology can be altered in highly trained individuals within days to weeks as compared to normal individuals. Studies also demonstrated that GM volume is greater in regions of the brain related to skill and expertise as shown in professional musicians [37] and for various athletes such as golfers [38], judokas [39], and divers [40].
On the other hand, Freud et al reported a 6% total reversible GM volume decline among athletes during a multi-step ultramarathon with a total distance of 4487 km in 64 days [34]. The decrease in GM volume was correlated to body weight loss in athletes during the race. Moreover, Freud argued that volume change in cerebral cortex could be explained by long-term exposure to stress such as hyponatremia, which is known to have a negative effect on brain structure [41,42]. However, the increase in cerebral GM volume seems to be unrelated to hyponatremia [43] and/or hypernatremia [44], since WM volume changes accompany GM volume changes during hyponatremia or hypernatremia as a result of serum osmolality changes [45]. The increase in total GM volume may be due to mild hypoxia as a result of total altitude change during UM races, but not marathon races (3370 m and 1120 m, respectively). However, Rupp et al [46] demonstrated higher GM and WM volumes after prolonged exercise compared to 10-h rest under hypoxic conditions. While changes in WM volume were correlated with arterial oxygen saturation and are explained as a result of hypoxia, GM volume remained the same during rest under hypoxic conditions. Rupp et al suggested that although it is not caused by hypoxemia, hypoxic exposure causes some disturbances of brain function such as increased GM volume. These studies together may suggest that endurance running has a greater impact on total GM compared to WM, and may provide a benefit in GM volume reserve, which is reported to decrease by 2% annually in sedentary males during aging [47,48].
Findings showed that hippocampus, basal ganglia, and thalamus components such as caudate and pallidus had greater volumes after the race in the UT group, whereas volumes of other brain structures were unchanged. These findings appear to agree with previous studies suggesting that some brain regions are more sensitive to exercise, especially ones that are prone to age-dependent atrophy [49,50]. Hippocampal and parahippocampal GM volume of the endurance athletes were significantly higher compared to performance athletes, which suggests that they cause changes in the motor learning and planning areas of the brain [51].
The present study showed that strenuous endurance exercise affects brain structures that were previously reported to be affected by aerobic exercise. In a cross-sectional study, it was shown that the higher the cardiorespiratory level, the larger the basal ganglia volume, which is related with better cognitive function performance [52]. Similarly, another study noted that caudate volume is closely related to the level of cardiorespiratory fitness and cognitive functions. Therefore, it can be predicted that an individual with higher cardiorespiratory fitness is more likely to have higher accuracy in task-switching [53]. The increase in cardiovascular fitness level and hippocampal volume due to exercise suggests that hippocampal volume plays a role in the fitness-spatial memory relationship [54]. A longitudinal study with 299 elderly subjects showed that greater physical activity above a threshold could double hippocampal volume in seniors [54]. Animal studies have demonstrated proliferation and survival of new neurons [55] and new vasculature [56] in the hippocampus after running exercise. Adult hippocampal neurogenesis was positively correlated to running distance and exercise duration in rats [57] with superior learning rates compared to sedentary animals [58].
Previous research showed that the caudate and putamen are activated differently during motor learning [59]. While the caudate usually plays a role in the beginning of motor learning, the putamen plays an active role in the automation of movements [60,61]. The globus pallidus, on the other hand, is a major component of the basal ganglia, which is responsible for the regulation of subconscious and voluntary movements [62]. Therefore, continuous changes in coordinative movements will result in some changes in cognitive functions [63] and might cause volumetric increase in some areas of the brain. The increased volume of caudate, pallidus, and thalamus in the current study might be explained by the requirement of adaptive tasks during outdoor running, which requires greater effort for balance and coordination compared to treadmill running.
The basal ganglia play a role in cognitive function through dopaminergic projections [64]. Therefore, it makes sense to assume that exercise causes upregulation of basal ganglia dopaminergic neurotransmission [65,66] and thereby improves cognitive processes after exercise. Interestingly, the fact that there was increased volume of the hippocampus, thalamus and basal ganglia in the UT group but not in the ST group indicates that there may be other factors in addition to exercise that affected brain-tissue volume alteration in this study. The UT group, but not the ST group, experienced sleep deprivation during the ultramarathon race, which lasted 24 hours. In 2019, Won et al showed that there was a relationship between executive functions and sleep and acute exercise, and that basal ganglia and caudate volumes are decisive in this interaction [67]. Studies have shown that sleep deprivation in rats decreased cell proliferation in the dentate gyrus of the hippocampus following a 96-hour treadmill exercise [68]. Findings of a similar study showed parallel results of neural effect [69]; however, the contribution of exercise volume and physical stress to neural effect has not been identified, since the effects of exercise and sleep deprivation are very similar. Conceptualizing the literature with the present results, it might be speculated that exercise amount and sleep deprivation in UT may produce different acute effects on the hippocampus, basal ganglia, and thalamus compared to ST, and further studies are needed to examine the interactive long- and short-term effects of sleep and endurance exercise on brain structure and function.
Although this study reports many valuable findings, it also has some limitations. First, we emphasize that our small sample was limited to healthy male older adult runners; thus, our findings may not be generalized to younger runners. Second, our findings are representative for single-stage ultramarathons at the distance of 120 km. Thus, findings cannot be generalized to all ultramarathon races because of their great diversity and additional extreme conditions such as weather, distance, and nature of the race. Moreover, the study was observational, and fluid and energy intake were not controlled during the international race. The restrictions of athletes from taking a shower right after the race and taking them to the MRI scan without any delay were tightly controlled to avoid any changes in the brain structures. Variables that may affect cortical thickness such as daytime sleepiness were not controlled in the observational study [70]; however, time-matched scanning procedure has been performed to lighten these alterations in brain volumes [71]. Although prior power analysis was performed, dropouts during the race caused a decline in the power of the study. Since evaluation of the acute effect of ultramarathon running on cortical and subcortical structures in the brain is sporadic, we compared our results with the current literature that is based on animal studies or long-term adaptations in human subjects. Both are insufficient to conclude specific outcomes, and further studies are needed to clarify the underlying mechanisms.
Conclusions
This study presented for the first time the acute effect of endurance running at 38.6 km and 119.8 km on the volume of cortical and subcortical brain structures by using high-resolution 3-Tesla MRI scanning. It can be concluded that single-stage endurance running event can cause increased cerebral GM volume in both short-trail and ultra-trail races. On the other hand, the increase in the volumes of the thalamus, caudate, pallidus, and hippocampus in the UT group, but not the ST group, might suggest that other variables such as sleep deprivation in addition to exercise may be crucial during UM running that extends beyond daylight hours into the night. It can be concluded that endurance running can help to preserve GM volume in male runners and suggests a method for prevention of stress or age-related GM decline. However, more detailed studies are needed to examine the cause of brain volume changes depending on both physical activity and physical stress experienced during ultramarathon running.
Figures
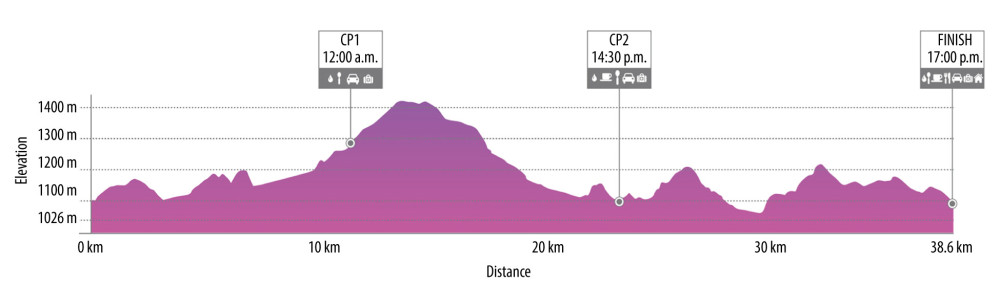 Figure 1. Schematic representation of 38.6-km short-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com).
Figure 1. Schematic representation of 38.6-km short-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com). 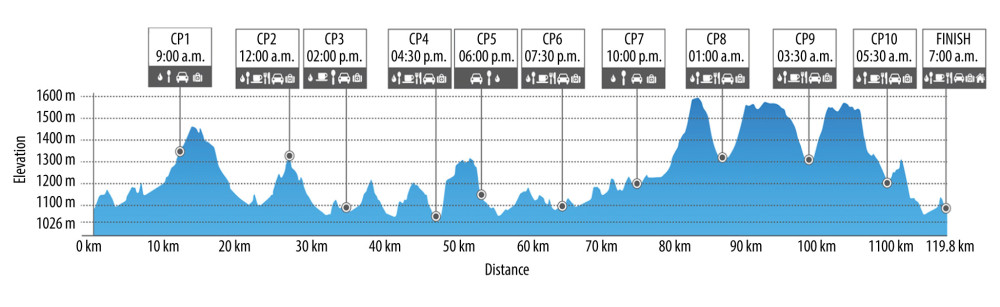 Figure 2. Schematic representation of 119.8-km ultra-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com).
Figure 2. Schematic representation of 119.8-km ultra-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com). 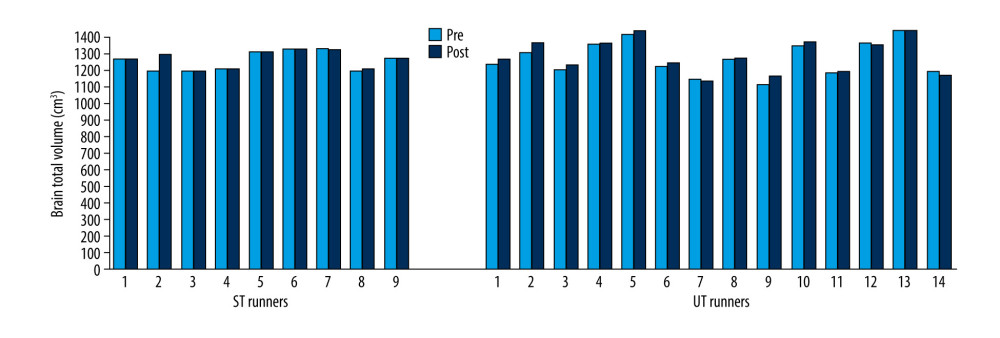 Figure 3. Brain total volume changes in ST and UT group for each runner.
Figure 3. Brain total volume changes in ST and UT group for each runner. 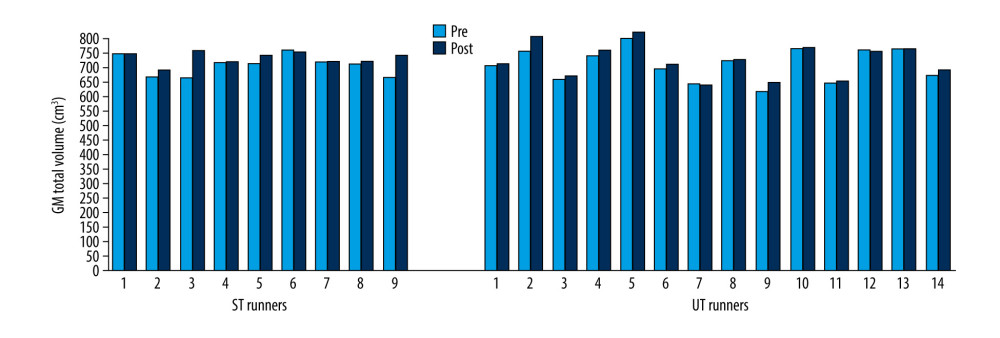 Figure 4. GM total volume changes in ST and UT group for each runner.
Figure 4. GM total volume changes in ST and UT group for each runner. 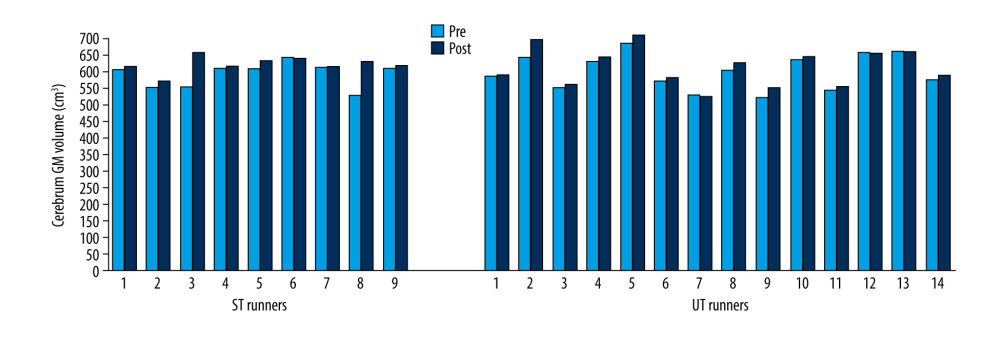 Figure 5. Cerebrum GM volume changes in ST and UT group for each runner.
Figure 5. Cerebrum GM volume changes in ST and UT group for each runner. 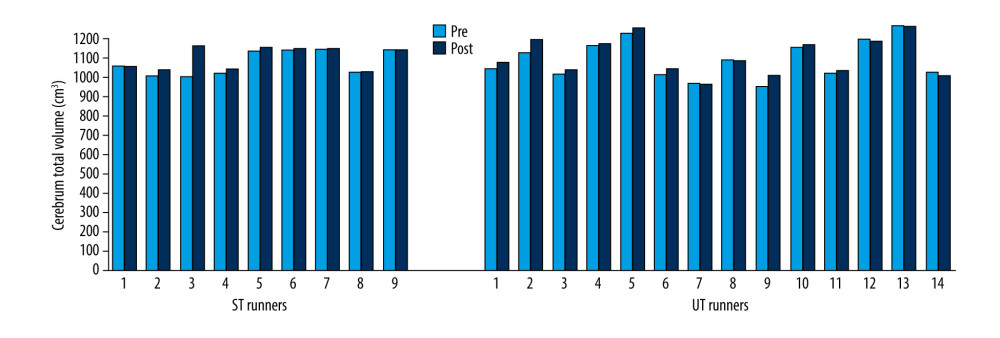 Figure 6. Cerebrum total volume changes in ST and UT group for each runner.
Figure 6. Cerebrum total volume changes in ST and UT group for each runner. 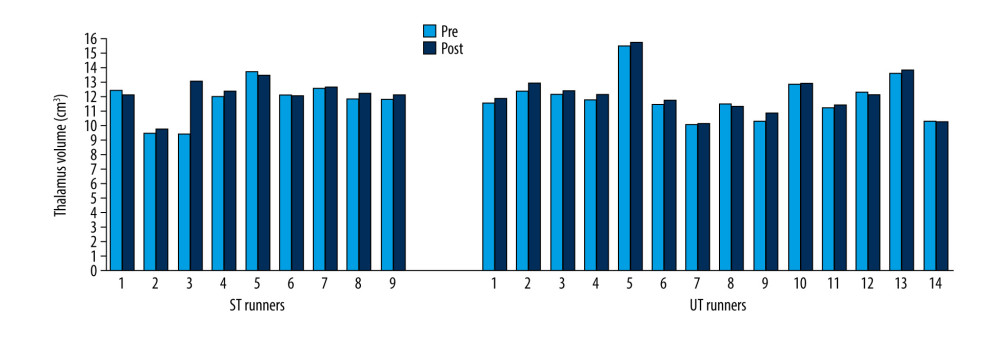 Figure 7. Thalamus volume changes in ST and UT group for each runner.
Figure 7. Thalamus volume changes in ST and UT group for each runner. 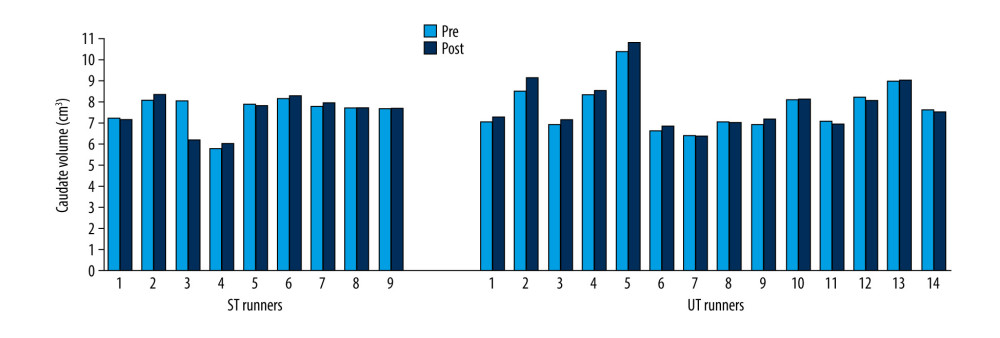 Figure 8. Caudate volume changes in ST and UT group for each runner.
Figure 8. Caudate volume changes in ST and UT group for each runner. 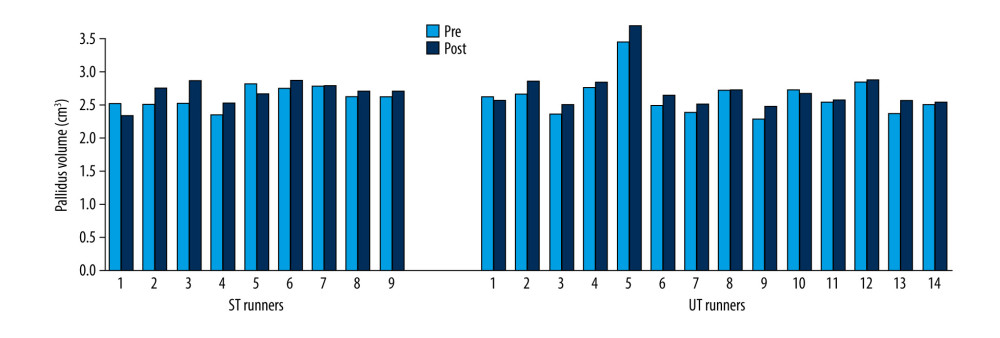 Figure 9. Pallidus volume changes in ST and UT group for each runner.
Figure 9. Pallidus volume changes in ST and UT group for each runner. 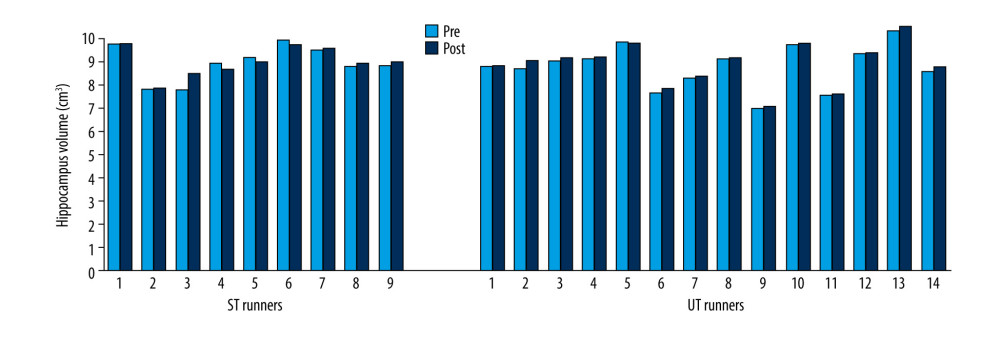 Figure 10. Hippocampus volume changes in ST and UT group for each runner.
Figure 10. Hippocampus volume changes in ST and UT group for each runner. Tables
Table 1. Descriptive characteristics of runners in ST and UT groups.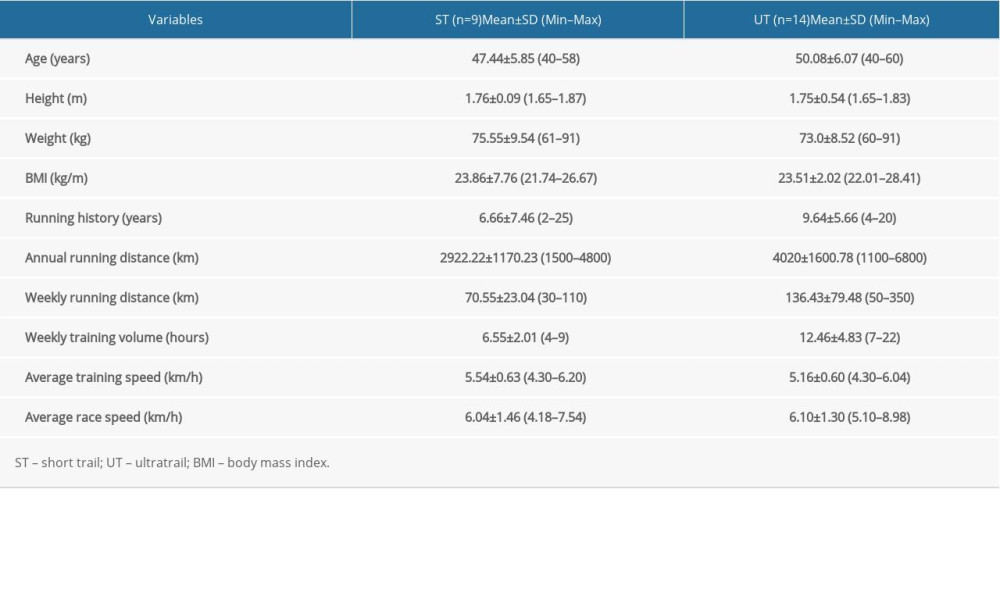 Table 2. Differences in volume of cortical and subcortical structures before and after 38.6 km race in short-trail group.
Table 2. Differences in volume of cortical and subcortical structures before and after 38.6 km race in short-trail group.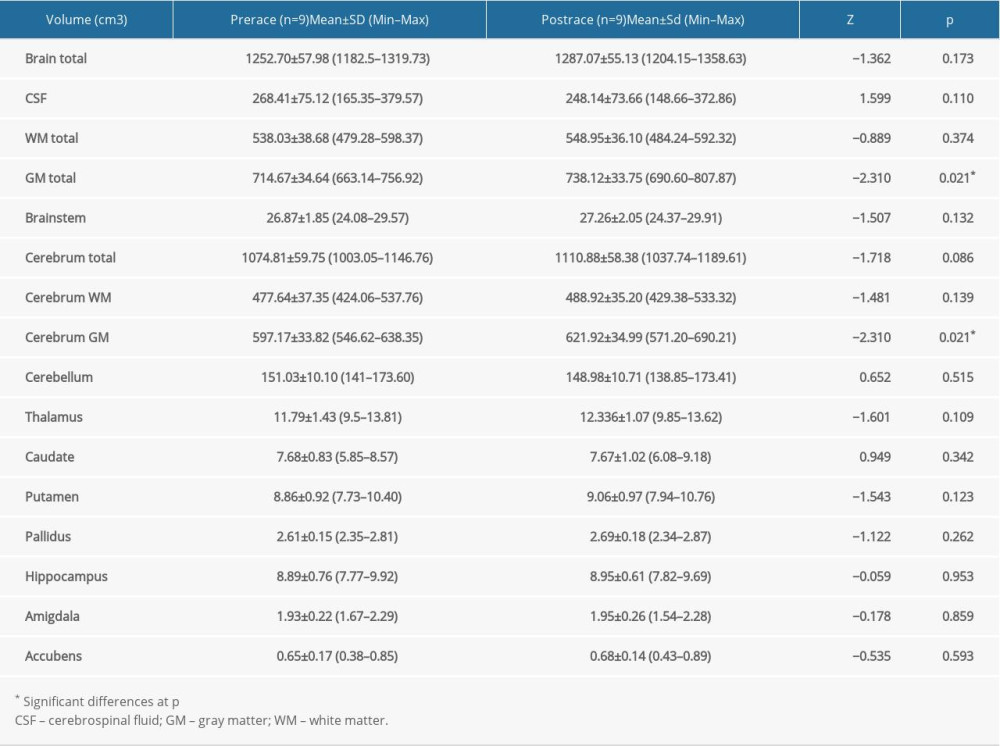 Table 3. Differences in volume of cortical and subcortical structures before and after 119.8 km race in ultra-trail group.
Table 3. Differences in volume of cortical and subcortical structures before and after 119.8 km race in ultra-trail group.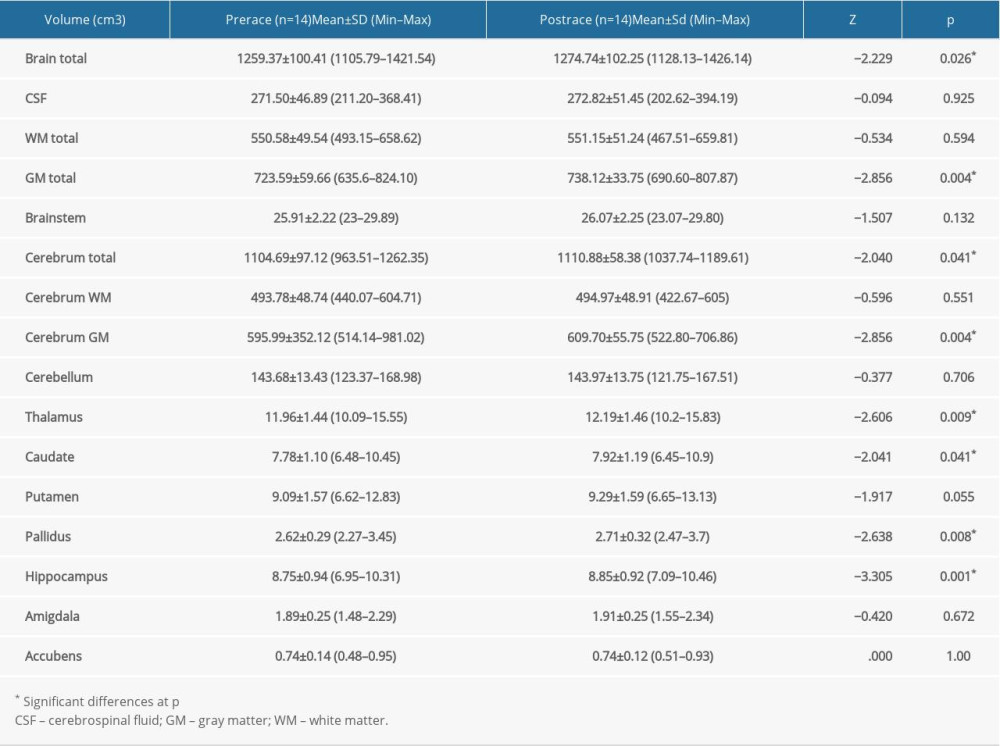
References
1. Garatachea N, Pareja-Galeano H, Sanchis-Gomar F, Exercise attenuates the major hallmarks of aging: Rejuvenation Res, 2015; 18(1); 57-89
2. Khoo J, Dhamodaran S, Chen DD, Exercise-induced weight loss is more effective than dieting for improving adipokine profile, insulin resistance, and inflammation in obese men: Int J Sport Nutr Exerc Metab, 2015; 25(6); 566-75
3. Mora S, Cook N, Buring JE, Physical activity and reduced risk of cardiovascular events: Potential mediating mechanisms: Circulation, 2007; 116(19); 2110-18
4. Wahl P, Mathes S, Köhler K, Acute metabolic, hormonal, and psychological responses to different endurance training protocols: Horm Metab Res, 2013; 45(11); 827-33
5. Sanchez LD, Corwell B, Berkoff D, Medical problems of marathon runners: Am J Emerg Med, 2006; 248(5); 608-15
6. Beutler J, Schmid E, Fischer S, Sudden cardiac death during a city marathon run: Der Anaesthesist, 2015; 64(6); 451-55
7. Fournier PE, Stalder J, Mermillod B, Chantraine A, Effects of a 110 km ultra-marathon race on plasma hormone levels: Int J Sports Med, 1997; 18(4); 252-56
8. McMurray RG, Forsythe WA, Mar MH, Hardy CJ, Exercise intensity-related responses of beta-endorphin and catecholamines: Med Sci Sports Exerc, 1987; 19(6); 570-74
9. Van Beaumont W, Strand JC, Petrofsky JS, Changes in total plasma content of electrolytes and proteins with maximal exercise: J Appl Physiol, 1973; 34(1); 102-6
10. Baur DA, Bach CW, Hyder WJ, Ormsbee MJ, Fluid retention, muscle damage, and altered body composition at the Ultraman triathlon: Eur J Appl Physiol, 2016; 116(3); 447-58
11. Kazimierczak M, Dabrowska A, Adamczewska K, Malchrowicz-Mosko E, The impact of modern ultramarathons on shaping the social identity of runners. The case study of Karkonosze winter ultramarathon: Int J Environ Res Public Health, 2019; 17(1); pii:E116
12. Shaw KL, Ostrow A, Motivation and psychological skills in senior athletes: Eur Rev Aging Phys Act, 2005; 2; 22-34
13. Spittler J, Oberle L, Current trends in ultramarathon running: Curr Sports Med Rep, 2019; 18(11); 387-93
14. Nikolaidis PT, Knechtle B, Age of peak performance in 50-km ultramarathoners – is it older than in marathoners?: Open Access J Sports Med, 2018; 9; 37-45
15. Esteve-Lanao J, Del Rosso S, Larumbe-Zabala E, Predicting recreational runners’ marathon performance time during their training preparation: J Strength Cond Res; 2019 [Epub ahead of print]
16. Waldvogel KJ, Nikolaidis PT, Di Gangi S, Women reduce the performance difference to men with increasing age in ultra-marathon running: Int J Environ Res Public Health, 2019; 16(13); 2377
17. O’Loughlin E, Nikolaidis PT, Rosemann T, Knechtle B, Different predictor variables for women and men in ultra-marathon running-the wellington urban ultramarathon 2018: Int J Environ Res Public Health, 2019; 16(10); 1844
18. Knechtle B, Rust CA, Knechtle P, Age-related changes in ultra-triathlon performances: Extrem Physiol Med, 2012; 1(1); 5
19. Knechtle B, Nikolaidis PT, Physiology and pathophysiology in ultra-marathon running: Front Physiol, 2018; 9; 634
20. Nikolaidis PT, Veniamakis E, Rosemann T, Knechtle B, Nutrition in ultra-endurance: State of the art nutrients: Nutrients, 2018; 10(12); 1995
21. Millet GP, Economy is not sacrificed in ultramarathon runners: J Appl Physiol (1985), 2012; 113(4); 686 author reply 687
22. Kim JH, Malhotra R, Chiampas G, Cardiac arrest during long-distance running races: N Engl J Med, 2012; 366(2); 130-40
23. Seal AD, Anastasiou CA, Skenderi KP, Incidence of hyponatremia during a continuous 246-km ultramarathon running race: Front Nutr, 2019; 6; 161
24. Krabak BJ, Lipman GS, Waite BL, Rundell SD, Exercise-associated hyponatremia, hypernatremia, and hydration status in multistage ultramarathons: Wilderness Environ Med, 2017; 28(4); 291-98
25. Vlachopoulos C, Kardara D, Anastasakis A, Arterial stiffness and wave reflections in marathon runners: Am J Hypertens, 2010; 23(9); 974-79
26. Hoppel F, Calabria E, Pesta D, Physiological and pathophysiological responses to ultramarathon running in non-elite runners: Front Physiol, 2019; 10; 1300
27. Colcombe SJ, Kramer AF, Erickson KI, Cardiovascular fitness, cortical plasticity, and aging: Proc Natl Acad Sci USA, 2004; 101(9); 3316-21
28. Burns JM, Cronk BB, Anderson HS, Cardiorespiratory fitness and brain atrophy in early Alzheimer disease: Neurology, 2008; 71(3); 210-16
29. Heyn PC, Johnson KE, Kramer AF, Endurance and strength training outcomes on cognitively impaired and cognitively intact older adults: A meta-analysis: J Nutr Health Aging, 2008; 12(6); 401-9
30. Erickson KI, Miller DL, Roecklein KA, The aging hippocampus. Interactions between exercise, depression, and BDNF: Neuroscientist, 2012; 18(1); 82-97
31. Liu PZ, Nusslock R, Exercise and hippocampal neurogenesis. A dogma re-examined and lessons learned: Neural Regen Res, 2018; 13(8); 1354-55
32. Wood KN, Nikolov R, Shoemaker JK: Front Aging Neurosci, 2016; 8; 155
33. Herm J, Haeusler KG, Kunze C, MRI brain changes after marathon running: Results of the berlin beat of running study: Int J Sports Med, 2012; 40(6); 856-62
34. Freund W, Faust S, Birklein F, Substantial and reversible brain gray matter reduction but no acute brain lesions in ultramarathon runners: Experience from the TransEurope-FootRace Project: BMC Med, 2012; 10(170); 1-11
35. Lohman TG, Roche AF, Martorell R: Anthropometric standardization reference manual, 1988, Human kinetics
36. Manjón JV, Coupé P, volBrain: An online MRI brain volumetry system: Front Neuroinform, 2016; 10; 30
37. Gaser C, Schlaug G, Brain structures differ between musicians and non-musicians: J Neurosci, 2003; 23(27); 9240-45
38. Jäncke L, Koeneke S, Hoppe A, The architecture of the golfer’s brain: PLoS One, 2009; 4(3); e4785
39. Jacini WF, Cannonieri GC, Fernandes PT, Can exercise shape your brain? Cortical differences associated with judo practice: J Sci Med Sport, 2009; 12(6); 688-90
40. Wei G, Zhang Y, Jiang T, Luo J, Increased cortical thickness in sports experts: A comparison of diving players with the controls: PLoS One, 2011; 6; e17112
41. Sapolsky RM, Glucocorticoids, stress, and their adverse neurological effects: Relevance to aging: Exp Gerontol, 1999; 34; 721-32
42. Landfield PW, Blalock EM, Chen KC, Porter NM, A new glucocorticoid hypothesis of brain aging: Implications for Alzheimer’s disease: Curr Alzheimer Res, 2012; 4(2); 205-12
43. Siegel AJ, Verbalis JG, Clement S, Hyponatremia in marathon runners due to inappropriate arginine vasopressin secretion: Am J Med, 2007; 120(5); 461.e11-17
44. Ayus JC, Varon J, Arieff AI, Hyponatremia, cerebral edema, and noncardiogenic pulmonary edema in marathon runners: Ann Intern Med, 2000; 132(9); 711-14
45. Biller A, Reuter M, Patenaude B: Am J Neuroradiol, 2015; 36(12); 2277-84
46. Rupp T, Jubeau M, Lamalle L, Cerebral volumetric changes induced by prolonged hypoxic exposure and whole-body exercise: J Cereb Blood Flow Metab, 2014; 34(11); 1802-9
47. Smith CD, Chebrolu H, Wekstein DR, Age and gender effects on human brain anatomy: A voxel-based morphometric study in healthy elderly: Neurobiol Aging, 2007; 28(7); 1075-87
48. Good CD, Johnsrude IS, Ashburner J, A voxel-based morphometric study of ageing in 465 normal adult human brains: Neuroimage, 2001; 14(1 Pt 1); 21-36
49. Colcombe S, Kramer AF, Fitness effects on the cognitive function of older adults: A meta-analytic study: Psychol Sci, 2003; 14(2); 125-30
50. Smith SM, Fox PT, Miller KL, Correspondence of the brain’s functional architecture during activation and rest: Proc Natl Acad Sci USA, 2009; 106(31); 13040-45
51. Schlaffke L, Lissek S, Lenz M, Sports and brain morphology – a voxel-based morphometry study with endurance athletes and martial artists: Neuroscience, 2014; 259; 35-42
52. Chaddock L, Erickson KI, Prakash RS, Basal ganglia volume is associated with aerobic fitness in preadolescent children: Dev Neurosci, 2010; 32(3); 249-56
53. Verstynen TD, Lynch B, Miller DL, Caudate nucleus volume mediates the link between cardiorespiratory fitness and cognitive flexibility in older adults: J Aging Res, 2012; 2012; 939285
54. Erickson KI, Raji CA, Lopez OL, Physical activity predicts gray matter volume in late adulthood: The Cardiovascular Health Study: Neurology, 2010; 75(16); 1415-22
55. van Praag H, Kempermann G, Gage FH, Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus: Nat Neurosci, 1999; 2(3); 266-70
56. Motl RW, Hubbard EA, Sreekumar N, Pallidal and caudate volumes correlate with walking function in multiple sclerosis: J Neurol Sci, 2015; 354(1–2); 33-36
57. Nokia MS, Lensu S, Ahtiainen JP, Physical exercise increases adult hippocampal neurogenesis in male rats provided it is aerobic and sustained: J Physiol, 2016; 594(7); 1855-73
58. Nichol KE, Parachikova AI, Cotman CW, Three weeks of running wheel exposure improves cognitive performance in the aged Tg2576 mouse: Behav Brain Res, 2007; 184(2); 124-32
59. Puttemans V, Wenderoth N, Swinnen SP, Changes in brain activation during the acquisition of a multifrequency bimanual coordination task: From the cognitive stage to advanced levels of automaticity: J Neurosci, 2005; 25(17); 4270-78
60. Steele CJ, Penhune VB, Specific increases within global decreases: A functional magnetic resonance imaging investigation of five days of motor sequence learning: J Neurosci, 2010; 30(24); 8332-41
61. Jueptner M, Weiller C, A review of differences between basal ganglia and cerebellar control of movements as revealed by functional imaging studies: Brain, 1998; 121(Pt 8); 1437-49
62. Gillies MJ, Hyam JA, Weiss AR, The cognitive role of the globus pallidus interna;Insights from disease states: Exp Brain Res, 2017; 235(5); 1455-65
63. Monno A, Temprado JJ, Zanone PG, Laurent M, The interplay of attention and bimanual coordination dynamics: Acta Psychologica, 2002; 110(2–3); 187-211
64. Crosson B, Benjamin M, Levy I: Role of the basal ganglia in language and semantics: Supporting cast, 2007; 219-43, New York, NY, USA, Cambridge University Press
65. Fisher BE, Petzinger GM, Nixon K, Exercise-induced behavioral recovery and neuroplasticity in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse basal ganglia: J Neurosci Res, 2004; 77(3); 378-90
66. Petzinger GM, Walsh JP, Akopian G, Effects of treadmill exercise on dopaminergic transmission in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury: J Neurosci, 2007; 27(20); 5291-300
67. Won J, Alfini AJ, Weiss LR, Caudate volume mediates the ınteraction between total sleep time and executive function after acute exercise in healthy older adults: Brain Plast, 2019; 5(1); 69-82
68. Guzmán-Marín R, Suntsova N, Stewart DR, Sleep deprivation reduces proliferation of cells in the dentate gyrus of the hippocampus in rats: J Physiol, 2003; 549(2); 563-71
69. Roman V, Van der Borght K, Leemburg SA, Sleep restriction by forced activity reduces hippocampal cell proliferation: Brain Res, 2005; 1065(1–2); 53-59
70. Killgore WD, Schwab ZJ, Kipman M, Voxel-based morphometric gray matter correlates of daytime sleepiness: Neurosci Lett, 2012; 518(1); 10-13
71. Carlson JM, Cha J, Fekete T, Left medial orbitofrontal cortex volume correlates with skydive-elicited euphoric experience: Brain Struct Funct, 2016; 221(8); 4269-79
Figures
Figure 1. Schematic representation of 38.6-km short-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com).Figure 2. Schematic representation of 119.8-km ultra-trail with detailed distance, altimetry, time zone, and check points (CP) (https://cappadociaultratrail.com).Figure 3. Brain total volume changes in ST and UT group for each runner.Figure 4. GM total volume changes in ST and UT group for each runner.Figure 5. Cerebrum GM volume changes in ST and UT group for each runner.Figure 6. Cerebrum total volume changes in ST and UT group for each runner.Figure 7. Thalamus volume changes in ST and UT group for each runner.Figure 8. Caudate volume changes in ST and UT group for each runner.Figure 9. Pallidus volume changes in ST and UT group for each runner.Figure 10. Hippocampus volume changes in ST and UT group for each runner. Tables
Table 1. Descriptive characteristics of runners in ST and UT groups.Table 2. Differences in volume of cortical and subcortical structures before and after 38.6 km race in short-trail group.Table 3. Differences in volume of cortical and subcortical structures before and after 119.8 km race in ultra-trail group.Table 1. Descriptive characteristics of runners in ST and UT groups.Table 2. Differences in volume of cortical and subcortical structures before and after 38.6 km race in short-trail group.Table 3. Differences in volume of cortical and subcortical structures before and after 119.8 km race in ultra-trail group. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387