12 August 2021: Review Articles
Recent Advances in Maturation of Pluripotent Stem Cell-Derived Cardiomyocytes Promoted by Mechanical Stretch
Xingwang Gu1ABCF, Fan Zhou2ABEF, Junsheng Mu3ADFG*DOI: 10.12659/MSM.931063
Med Sci Monit 2021; 27:e931063






Abstract
ABSTRACT: Stem cells have significant potential use in tissue regeneration, especially for treating cardiac diseases because of their multi-directional differentiation capability. By mimicking the in vivo physiological environment of native cardiomyocytes during their development and maturation, researchers have been able to induce pluripotent stem cell-derived cardiomyocytes (PSC-CMs) at high purity. However, the phenotype of these PSC-CMs is immature compared with that of adult cardiomyocytes. Various strategies have been explored to improve the maturity of PSC-CMs, such as long-term culturing, mechanical stimuli, chemical stimuli, and combinations of these strategies. Among these strategies, mechanical stretch as a key mechanical stimulus plays an important role in PSC-CM maturation. In this review, the optimal parameters of mechanical stretch, the effects of mechanical stretch on maturation of PSC-CMs, underlying molecular mechanisms as well as existing problems are discussed. Mechanical stretch is a powerful approach to promote the maturation of SC-CMs in terms of morphology, structure, and functionality. Nonetheless, further research efforts are needed to reach a satisfactory standard for clinical applications of PSC-CMs in treating cardiac diseases.
Keywords: Engineering, Mechanical Processes, Pluripotent Stem Cells, Stress, Mechanical
Background
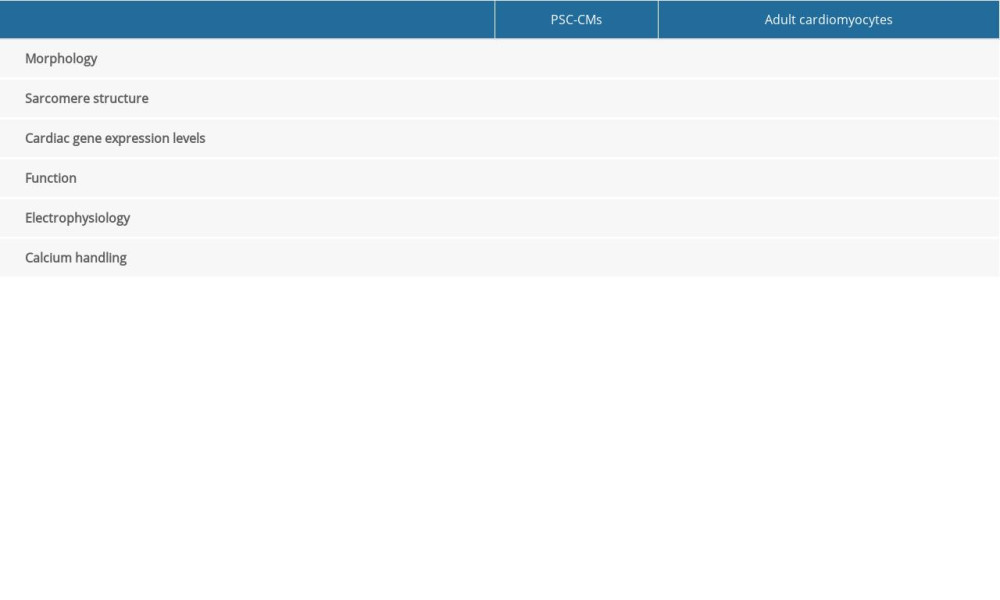
For patients with cardiac diseases like myocardial infarction and heart failure, many strategies have been tried to maintain and even reverse the impaired cardiac function in the long term. Among them, cell therapy holds significant promise as it provides the heart with new cells to overcome the fundamental cause of these diseases, the loss of cardiomyocytes [1]. Techniques such as engineered heart tissue (EHT) [2] are also used to avoid the shortcomings existing with early attempts to provide cells through intramyocardial and intravenous injection, like low survival rate and low retention rate of cells at the impaired area [3,4]. In terms of available cell sources, stem cells are generally a better choice than differentiated cells because the latter usually cannot re-differentiate to qualified cardiomyocytes and cannot establish good electrical integration with the host cardiac tissue, thus having limited curative effects [5,6]. Both pluripotent stem cells (PSCs) and adult stem cells, like bone marrow mesenchymal stem cells, are capable of differentiating to cardiomyocytes [7]. PSCs are primarily embryonic stem cells (ESCs) that are derived from an inner-cell mass of the preimplanted blastocyst or from primordial germ cells of the early embryos, and induced pluripotent stem cells (iPSCs) that are derived from somatic cells by using reprogramming factors and resemble ESCs in many ways. Although use of these cells faces ethical and safety concerns [8], they are widely studied for their superior differentiation potential over that of adult stem cells, and can hopefully be used for various therapeutic purposes in regenerative medicine in the future [9]. However, as early work tried to obtain cardiomyocytes from PSCs via different methods, such as the use of exogenous cytokines, growth factors, and extracellular matrix [10], these pluripotent stem cell-derived cardiomyocytes (PSC-CMs) turned out to be significantly more immature compared to adult cardiomyocytes and exhibit a fetal-like phenotype [11]. For example, PSC-CMs are generally round and single-nucleated, while adult cardiomyocytes are elongated, rod-shaped, and partially binucleated. Other differences between them are briefly summarized in Table 1 [12]. These major shortcomings limited their applications for treating cardiac diseases. Therefore, finding approaches to improve the PSC-CM maturation process remains a key issue to resolve.
Recent strategies have been explored to improve PSC-CM maturation, such as long-term culturing, substrate stiffness, electric stimulation, biochemical cues, and mechanical stimuli [13]. In vivo, the pulsating heart is constantly subjected to the preload caused by filling of the heart cavity by the returning blood flow. As mechanical stimuli may be critical to cardiac development [14], applying mechanical stretch that simulates the normal myocardium preload may effectively promote the maturation of PSC-CMs. As early as 2000, Zimmermann et al found that neonatal rat cardiomyocytes-based EHT retained many of the physiological characteristics of rat cardiac tissue [15]. After phasic mechanical stretching (also termed mechanical strain), cardiac cells in EHT were extensively interconnected and longitudinally oriented. Cardiac muscle bundles also showed morphological features that resembled adult rather than immature native tissue [16], which revealed the potential value of mechanical stretch in the maturation of immature cardiomyocytes. In a subsequent study by Shimko et al, a custom-built device was used to exert mechanical stretch to murine ESC-CMs (mESC-CMs) embedded in collagen and fibronectin scaffolds. These mESC-CMs showed longitudinal alignment within the constructs with long strands of cells containing elongated nuclei. Mechanically loaded constructs also presented thin interwoven strands between the cells, similar to that observed in the neonatal heart. Changes in the expression of particular cardiac genes such as α-skeletal actin, α-cardiac actin, α-myosin heavy chain (MHC), and connexin-43 (Cx-43) were also observed [17]. These results further proved the role of such stimulation in the maturity of PSC-CMs. However, despite these impressive advances in the use of mechanical stimuli to promote PSC-CM maturation, there is no recent review focusing on this topic. In this review, we summarize the parameters of applying mechanical stretch as well as its impact on the maturation of PSC-CMs. Then, the underlying signal transduction pathways and existing problems are discussed.
Parameters of Mechanical Stretch
(1) STRETCH DIRECTION:
Uniaxial stretch is the primary parameter adjusted for mechanical stretching of engineered tissue [17,22,23] and is dependent on the design of the bioreactors. Various axial directions can be used when trying to differentiate stem cells into SC-CMs by mechanical stretch, including biaxial, equiaxial, and multiaxial stretching [24–26]; however, they are rarely used when trying to improve the maturity of PSC-CMs [20]. Currently, no studies have explored differences caused by these axial directions.
(2) STRETCH INTENSITY:
Although data examining stretch intensity in defining the optimal degree of elongation are sparse, setting the stretch intensity to around 10% elongation ensures an efficient improvement in human PSC-CM (hPSC-CMs) maturation [19,22,27]. When Kroll et al applied a 5% cyclic stretch to human iPSC-CMs (hiPSC-CMs) cultured on PDMS membranes, these cells did not mature to cardiac phenotypes in morphology and cardiac marker expression levels [28]. In another study, 15% stretch applied to hESC-CMs caused a significant increase in cell volume and sarcomere size but was also associated with a 1.6-fold slower beating rate. This abnormal beating rate was caused by the greater downregulation of Ca2+, Na+, and K+ channel proteins [20]. Considering that 10%–20% elongation may be physiological [29,30], these studies may provide an approximate range of optimal stretch amplitude for hPSC-CMs. Further studies are needed to confirm these observations.
(3) STRETCH FREQUENCY:
As for stretch frequency, cyclic stretching may have a better effect than static stretching (0 Hz) [27,31], and the majority of studies based on hPSC-CMs have selected 1 Hz to simulate the human heart rate [20,32]. For mouse models, a study based on mouse ESC-CMs (mESC-CMs) compared the different effects of 1, 2, and 3 Hz stretching at 10% elongation [17]. Only the 3 Hz stretch significantly upregulated cardiac genes, such as α-skeletal actin, α-cardiac actin, and α-MHC, which indicates that a higher frequency may better mimic the murine heart rate, a point also raised in other reports [33]. In contrast, a recent experiment showed cyclic stretch conditions (2.5%, 5%, and 10% strain) at 0.33 Hz on mESC-CMs were also able to significantly increase the expression of cardiac genes [34]. This controversy may be partly due to the different study designs and different experimental conditions used, including the longer duration of cell culture in the latter study. In addition, because a normal heart does not always keep the same beating rate, it is noteworthy that a dynamic change in stretch frequency may also have some effects on cardiomyocyte maturation [27].
(4) STRETCH TIME:
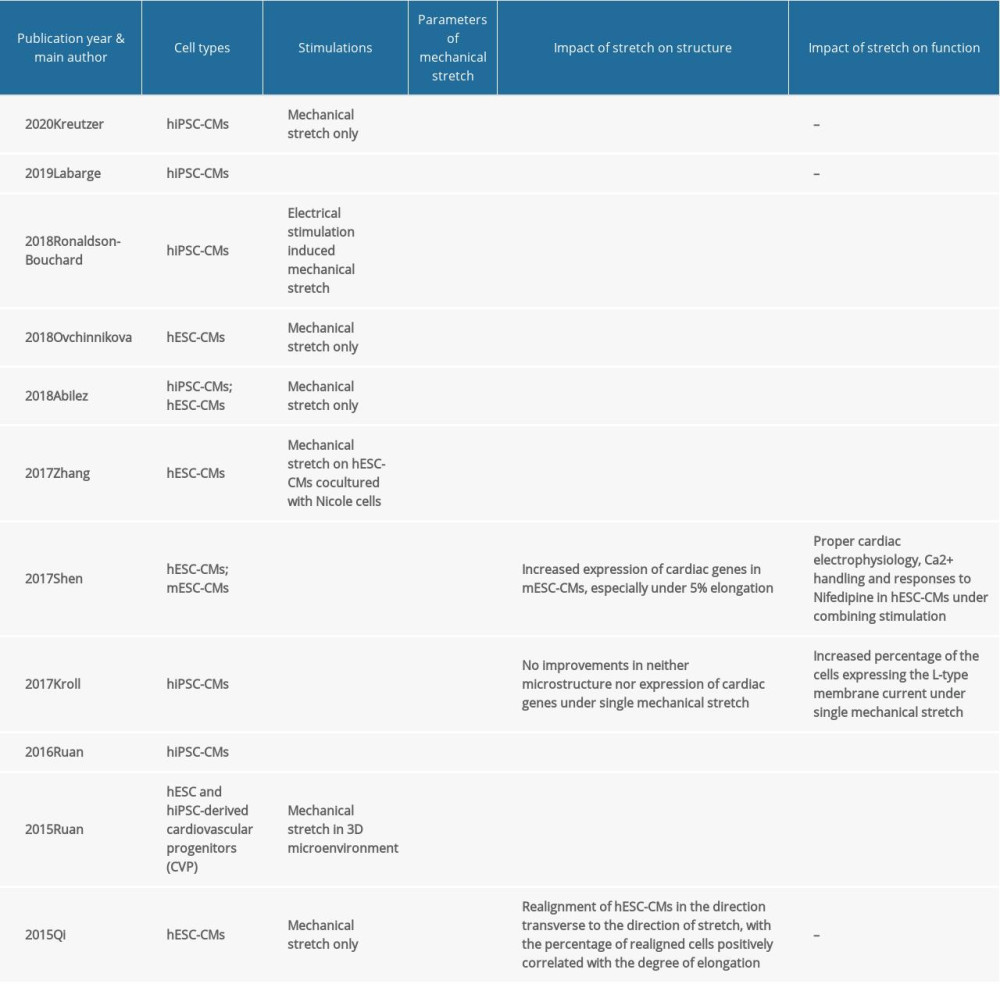
As long-term culturing advances maturity of PSC-CMs [35], application of longer stretch time should yield better results if carried out under an appropriate stretch regimen. Typically, the stretch time is selected to be no more than 2 weeks, as is shown in Table 2. Considering that excessive stretch time may increase the time and economic burden of its future clinical application, more research is required to explore the balance in between. Importantly, another study revealed that mechanical conditioning should be initiated early during the high cell plasticity period because the responsiveness of hiPSC-CMs to physical stimuli declines as maturation progresses [36].
Effects of Mechanical Stretch on Maturation of PSC-CMs
Structural Level
STRUCTURAL LEVEL:
During the development of cardiomyocytes, the changes in cell morphology, alignment, and microstructures (such as sarcomere, T-tubes, and mitochondria) lay the foundation for future mature contractility and electrophysiological performance. In addition, evaluating their cardiac gene expression profiles is also a powerful tool to determine the maturation stage of PSC-CMs. In this review, the description of structural changes to PSC-CMs under mechanical stretch is divided into 2 parts: structures observable by imaging techniques, and cardiac gene expression levels.
Tulloch et al [37] examined, through imaging, the influence of static and cyclic stretching (5% elongation, 4 days of 1 Hz) on intercellular alignment of hiPSC-CMs and hESC-CMs, which was quantified from the reciprocal of the cell axis angle dispersion, in which a low standard deviation of angles indicated a high degree of alignment. Both static stretch and cyclic stretch of cells increased the alignment value (4.09 for both stretch groups vs 1.96 for the non-stretch group). Orientation analysis with circular variance of hiPSC-CMs receiving a uniaxial (8%, 0.8 Hz) stretch on PDMS membranes for 7 days clearly demonstrated a significant unidirectionality of sarcomere structures that were perpendicular to the stretch axis after 2 days, while sarcomeres were only slightly more oriented at days 4 and 7 [23]. The authors of that study concluded that a time span of 4 days was sufficient to gain the highest degree of orientation. hiPSC-CMs in another study [22] formed many Z-bodies, the precursors to Z-bands, adjacent to contractile fibers under uniaxial (10%, 1 Hz) stretch for 7 days. Interestingly, mechanical stretch has been shown to cause rearrangement of hESC-CMs and human atrial fibroblasts perpendicular to the direction of the stretch [32,38], whereas another study revealed parallel rearrangement of newborn mouse cardiomyocytes [39]. Furthermore, in a long-term study, hPSC-CMs cultured under static stretch showed heterogeneous cellular phenotypes, with ventricular-like cells being the predominant phenotype (57%) along with atrial-like (24%) and nodal-like (19%) cells, when the culture time was extended to 25–28 days [40]. Other experiments focusing on hESC-CMs showed similar results in terms of cell morphology and sarcomere structure [20,27,32], which indicates that improved cardiac maturity benefited from mechanical stretch. Apart from stretch-only conditions, Kensah et al investigated the combined role of fibroblasts, ascorbic acid, and mechanical stimuli on both human and murine PSC-CMs, showing that these cells presented even better organization compared with that of a single stimulus [41]. However, despite all these impressive outcomes, the alignment level of such stimulated hPSC-CMs still cannot reach that observed in neonatal or adult rat cardiomyocytes [37]. Although it is inappropriate to compare cells from 2 species, this observation in rats highlights the differences between stretched PSC-CMs and mature cardiomyocytes.
Genes regulated by mechanical stretch within PSC-CMs are related to contractility (eg, cTnI, α-MHC and MLC2v), gap junctions (eg, Cx-43 and N-cadherin), ion channels (eg, SCN9A), and others (eg, ANP) [20,27. Although many genes have been detected by various studies, only those genes that are studied frequently are discussed in this review.
(1) MYOSIN HEAVY CHAIN (MHC): In humans, the MHC isoforms MHC-α and MHC-β change with the ratio of MHC-β over MHC-α, increasing as cardiac development matures [42]. Mechanical stretch (1 Hz, 5% elongation for 2 weeks) on hESC-CMs has been shown to increase the expression of MHC-β by 550% to 800% compared with hESC-CMs that experienced non-stretch or static stretch conditions [31]. Concurrently, MHC-α gene expression in hESC-CMs decreased by 62% or 50% under cyclic stretch conditions compared with MHC-α gene expression in cells under non-stretch or static stretch conditions, respectively. Similar increases in the MHC-β/MHC-α ratio by mechanical stretch were observed in other studies examining hiPSC-CMs and hESC-CMs [21,22]. It is noteworthy that in mouse models, although MHC-α is the predominate MHC isoform in the adult heart, this only occurs after the mouse is born. During pregnancy, the amount of MHC-β is still higher and the MHC-β/MHC-α ratio is even progressively increased [43,44]. This special developing pattern may help better evaluate the maturation status of mPSC-CMs under mechanical stretch.
(2) CONNEXIN 43 (CX-43): Cx-43 is the main gap junction protein expressed in the heart. It has been shown to gradually accumulate during gestational stage in rats [45]. For the mouse and human fetal heart, the increase of Cx-43 during gestation is associated with the development of the right ventricle and ventricular conduction systems, respectively [46–48]. In a study where stretch was applied (10% elongation, 3 Hz) to mESC-CMs cultured in collagen gels, Cx-43 was observed to be more widespread and diffuse throughout the tissue compared with that of the non-stretch group [17]. For mhiPSC-CMs, stretch (10% elongation, 1 Hz) for 7 days also induced the expression of Cx-43 [41]; however, the levels were up to 10-fold lower compared with that of neonatal hearts. These results showed that mechanical stretch accelerates the accumulation of Cx-43, indicating the advanced maturity of mPSC-CMs, while the effects may be still far lag behind naturally developed heart cells. Similar results were also found in hESC-CMs and hiPSC-CMs [22,31].
FUNCTIONAL LEVEL:
The main features used to assess the function of PSC-CMs are active force (twitch force), passive force (stiffness, elastic modulus), and ion dynamics. Ruan et al found hESC-CMs in collagen hydrogels under uniaxial stretch (1 Hz, 5% elongation) presented significantly improved active and passive forces. A non-stretch construct had a passive stiffness of 0.22±0.04 kPa, whereas hESC-CMs in collagen hydrogels subjected to static and cyclic stretching increased passive stiffness to 0.47±0.05 kPa and 0.71±0.12 kPa, respectively [31]. Kensah et al explored the combined impact of ascorbic acid and growing static stretch (stepwise elongation every second day) on human and murine PSC-CMs [41]. For miPSC-CMs, maximum active forces almost doubled, measured at 1.42±0.09 mN/mm2, whereas passive forces were about 3 times higher than in untreated controls at day 21. Similar results were found in hPSC-CMs. In addition, the force-length relationship can be evaluated by installing a force transducer on the mechanical arm where the tissue is fixed while gradually stretching the cells. A linear relationship was demonstrated for hESC-CMs subjected to uniaxial (1 Hz, 10%) stretch for 3 days, which is in agreement with the Frank-Starling Law because native myocardia behave in the same way [27]. However, the hESC-CMs in this study did not show a positive force-frequency relationship, which is observed in native myocardia, because their force output declined as the contraction rate accelerated. In another study, mechanical stretch was observed to weaken the original negative correlation of hPSC-CMs and promote the development of a positive correlation [49]. Importantly, if these tests need separation between cells and their culture environment, the outcomes may incorrectly reflect the in situ states of PSC-CMs. Some specific devices have been developed to realize in situ monitoring of cellular functions to solve this problem [50,51]. Moreover, for some drugs such as isoproterenol, hESC-CMs respond to the drugs in a similar manner to native cardiomyocytes, with corresponding increases or decreases in contractile force observed [31,52]. These enhancements may be attributed to the development of the intracellular sarcomere, mitochondria [18], and realignment of cells [39].
In terms of ion dynamics, hiPSC-CMs express the main components of cardiac excitation-contraction coupling, including L-type calcium channels and sodium-calcium exchangers [53–56]. hiPSC-CMs also generate ion currents for depolarization (INa, ICaL, If) and repolarization (Ito, IKr, IKs, IK1) of the membrane, which together produce action potential waveforms resembling that of human cardiomyocytes [57]. However, the poor co-localization of calcium channels and ryanodine receptors as well as the non-uniform distribution of calcium release [58,59] caused the upstroke and decline rates of the whole-cell Ca2+ signals to be substantially slower in hiPSC-CMs compared with that observed in adult cardiomyocytes [60,61]. These findings reveal the gap in ion dynamics, especially Ca2+ handling, between hiPSC-CMs and adult cardiomyocytes. A previously mentioned study found that with uniaxial cyclic strain at 1 Hz, 5% elongation on hESC-CMs was able to increase both peak calcium flux and upstroke rate by over 100% [31]. A study by Kroll et al [28], applying the same set of stretch parameters on hiPSC-CMs, also showed an increase in the percentage of cells expressing an L-type membrane current (from 66% to 81%) compared with that of the non-stretch group. Similar to contractility, proper responses of Ca2+ handling to cardiac drugs such as Nifedipine were also observed [34]. Together, these changes in parallel with the expression levels of ion channels [20,21] indicate the improved maturity of PSC-CMs under mechanical stretch in vitro.
Signal Transduction Mechanisms
(1) TRPV4/PI3K/AKT:
The TRPV4 channel is a non-selective cation channel that is Ca2+ permeable and activation of this channel causes an increase in intracellular Ca2+ levels [62]. Qi et al demonstrated that uniaxial cyclic stretching induces an increase Ca2+ influx, AKT phosphorylation, and realignment of hESC-CMs, and all these effects are abolished by TRPV4 inhibitors. Additionally, inhibitors of AKT and PI3K abolished stretch-induced hESC-CM realignment, suggesting the possible role of PI3K/AKT in cell realignment under cyclic stretch [32]. Because TRPV4 should be upstream of AKT, and TRPV4 channels are Ca2+ permeable channels whose activity leads to a rise in the intracellular Ca2+ level, it is likely that Ca2+ is a link between TRPV4 and AKT activity, which has been shown in other studies [63,64]. These studies show that the TRPV4/PI3K/AKT signal pathway may mediate the realignment of hESC-CMs under cyclic stretch.
(2) WNT/β-CATENIN:
The Wnt/β-catenin signal pathway is well known to influence a diverse set of biological processes, especially those processes involved in embryonic development and tissue renewal [65]. Recently, the Wnt/β-catenin signal pathway has been demonstrated to play a biphasic role in cardiogenesis (positive, then negative as the heart develops) [66,67]. Shen et al concluded that after 20 days of cyclic stretch, hESC-CMs expressed a lower level of β-catenin compared with that of static hESC-CMs, which indicates an inhibition of the Wnt/β-catenin signal pathway because of mechanical forces [34]. In their study, hESC-CMs under cyclic stretch displayed advanced maturity of the sarcomere structure and expression of cardiac markers such as Cx-43 and cTnT compared with those for hESC-CMs under static conditions. However, neither the development stage of stretched hESC-CMs nor changes caused by mechanical stretch through the Wnt/β-catenin signal pathway were explored. Further studies are required to characterize the vital role of the Wnt/β-catenin signal pathway in late cardiogenesis.
(3) OTHERS:
In embryonic mouse cardiomyocytes, stretch decreases the expression of the TGF-β family of proteins and the phosphorylation level of SMAD3, which is located downstream of TGF-β [68]. In bone marrow mesenchymal stem cells, cyclic stretching promoted these stem cells to differentiate into cardiomyocyte-like cells by suppressing the expression of the miR-27 and SCF genes (a target of miR-27). Although these findings did not originate from studies examining PSC-CMs, they may provide new clues for exploring potential molecular mechanisms regulated by mechanical stretch during the maturation of PSC-CMs.
Discussion and Summary
As is discussed in the “Parameters of mechanical stretch” section, even though there has been a growing number of studies based on PSC-CMs in recent years, few of them are focused on finding the appropriate stretch parameters. A noticeable issue is the lack of versatile and universally used devices that release/apply proper mechanical stretch to meet most requirements of different researchers. Another problem is the diversity of study designs, including different stem cell sources, dimensions of extracellular microenvironment (2D or 3D), additional stimuli like electrical stimulation, as well as diverse indicators for assessing the maturation status of PSC-CMs, leading to difficulty in comparing different trials. These shortcomings are partially due to the fact that mechanical stretch alone is not enough to fully promote the maturity of PSC-CMs, an issue that will be addressed later. Based on current progresses, suggestions on developing more standardized devices and study designs are as follow: (1) Since native myocardium is also stretched from different directions and even different planes (like a small part of the surface of an inflating balloon), devices which are pneumatically or vacuum controlled [69] to make the tissue extend nonlinearly may have a better effect than flexible pillars that stretch the tissue in a single straight direction; (2) 3D extracellular matrix should be considered, as it not only closely mimic the native environment where cardiomyocytes are subjected to mechanical stretch, but also can promote the maturity of PSC-CMs alone [70]; (3) In addition to those specially designed research purposes, some indicators that are easy to quantify should be universally proposed for maturation assessment, such as sarcomere length, expression levels of specific cardiac genes, active contracting force, and passive stiffness of the tissue; (4) In particular, comparing stretched PSC-CMs to adult cardiomyocytes in each study may be vital in identifying the real maturation stage of the former.
Indeed, mechanical stretch is able to promote the maturity of PSC-CMs to some extent, including increased size and elongated alignment of cells and sarcomeres, improved expression of cardiac markers (eg, actin, MHC-β, Cx-43, MLC2v), stronger contractility and passive stiffness, enhanced calcium handling, as well as ability to respond to some medicines in a way similar to that of adult cardiomyocytes. However, as discussed, the maturity of PSC-CMs under stretch conditioning still cannot reach a satisfactory level for further clinical usage. This is consistent with other studies where PSC-CMs generally remain within the early to late fetal cardiomyocyte stages after applying engineering approaches like electrical stimulation, non-cardiomyocyte interactions, or extracellular matrix interactions [12]. Considering that native heart muscle cells require a long time to mature in a complex physiological environment by receiving various stimuli, these outcomes under single in vitro simulated condition within a restricted culture time seem to be reasonable. Although researchers have been exploring the underlying signal transduction pathway and the combination of different maturation-promoting strategies [71], the number of relevant studies is limited. Further efforts should be focused on these aspects as well. Hopefully, with advances in research examining the effects of mechanical stretch on the maturation of PSC-CMs, we may soon develop more effective stem cell therapies for treating cardiac diseases.
References
1. Lin Z, Pu WT, Strategies for cardiac regeneration and repair: Sci Transl Med, 2014; 6(239); 239rv231
2. Hirt MN, Hansen A, Eschenhagen T, Cardiac tissue engineering: State of the art: Circ Res, 2014; 114(2); 354-67
3. Geng YJ, Molecular mechanisms for cardiovascular stem cell apoptosis and growth in the hearts with atherosclerotic coronary disease and ischemic heart failure: Ann NY Acad Sci, 2003; 1010; 687-97
4. Poynter JA, Herrmann JL, Manukyan MC, Intracoronary mesenchymal stem cells promote postischemic myocardial functional recovery, decrease inflammation, and reduce apoptosis via a signal transducer and activator of transcription 3 mechanism: J Am Coll Surg, 2011; 213(2); 253-60
5. Sanganalmath SK, Bolli R, Cell therapy for heart failure: A comprehensive overview of experimental and clinical studies, current challenges, and future directions: Circ Res, 2013; 113(6); 810-34
6. Segers VFM, Lee RT, Stem-cell therapy for cardiac disease: Nature, 2008; 451(7181); 937-42
7. Krishna KA, Krishna KS, Berrocal R, Myocardial infarction and stem cells: J Pharm Bioallied Sci, 2011; 3(2); 182-88
8. Volarevic V, Markovic BS, Gazdic M, Ethical and safety issues of stem cell-based therapy: Int J Med Sci, 2018; 15(1); 36-45
9. Pushp P, Nogueira DES, Rodrigues CAV, A concise review on induced pluripotent stem cell-derived cardiomyocytes for personalized regenerative medicine: Stem Cell Rev Rep, 2020 [Online ahead of print]
10. Heng BC, Haider H, Sim EK, Strategies for directing the differentiation of stem cells into the cardiomyogenic lineage in vitro: Cardiovasc Res, 2004; 62(1); 34-42
11. Feric NT, Radisic M, Maturing human pluripotent stem cell-derived cardiomyocytes in human engineered cardiac tissues: Adv Drug Deliv Rev, 2016; 96; 110-34
12. Scuderi GJ, Butcher J, Naturally engineered maturation of cardiomyocytes: Front Cell Dev Biol, 2017; 5; 50
13. Yang X, Pabon L, Murry CE, Engineering adolescence maturation of human pluripotent stem cell-derived cardiomyocytes: Circ Res, 2014; 114(3); 511-23
14. Nguyen M-D, Tinney JP, Ye F, Effects of physiologic mechanical stimulation on embryonic chick cardiomyocytes using a microfluidic cardiac cell culture model: Anal Chem, 2015; 87(4); 2107-13
15. Zimmermann WH, Fink C, Kralisch D, Three-dimensional engineered heart tissue from neonatal rat cardiac myocytes: Biotechnol Bioeng, 2000; 68(1); 106-14
16. Zimmermann WH, Schneiderbanger K, Schubert P, Tissue engineering of a differentiated cardiac muscle construct: Circ Res, 2002; 90(2); 223-30
17. Shimko VF, Claycomb WC, Effect of mechanical loading on three-dimensional cultures of embryonic stem cell-derived cardiomyocytes: Tissue Eng Part A, 2008; 14(1); 49-58
18. Kim HK, Kang YG, Jeong SH, Cyclic stretch increases mitochondrial biogenesis in a cardiac cell line: Biochem Biophys Res Commun, 2018; 505(3); 768-74
19. Llucià-Valldeperas A, Bragós R, Soler-Botija C, Unravelling the effects of mechanical physiological conditioning on cardiac adipose tissue-derived progenitor cells in vitro and in silico: Sci Rep, 2018; 8(1); 499
20. Ovchinnikova E, Hoes M, Ustyantsev K, Modeling human cardiac hypertrophy in stem cell-derived cardiomyocytes: Stem Cell Rep, 2018; 10(3); 794-807
21. Mihic A, Li J, Miyagi Y, The effect of cyclic stretch on maturation and 3d tissue formation of human embryonic stem cell-derived cardiomyocytes: Biomaterials, 2014; 35(9); 2798-808
22. Labarge W, Mattappally S, Kannappan R, Maturation of three-dimensional, hipsc-derived cardiomyocyte spheroids utilizing cyclic, uniaxial stretch and electrical stimulation: PLoS One, 2019; 14(7); e0219442
23. Kreutzer J, Viehrig M, Pölönen R-P, Pneumatic unidirectional cell stretching device for mechanobiological studies of cardiomyocytes: Biomech Model Mechanobiol, 2020; 19(1); 291-303
24. Girão-Silva T, Bassaneze V, Campos LC, Short-term mechanical stretch fails to differentiate human adipose-derived stem cells into cardiovascular cell phenotypes: Biomed Eng Online, 2014; 13; 54
25. Abd Emami B, Mahmoudi E, Shokrgozar MA, Mechanical and chemical predifferentiation of mesenchymal stem cells into cardiomyocytes and their effectiveness on acute myocardial infarction: Artif Organs, 2018; 42(6); E114-26
26. Cao C, Li L, Li H, Cyclic biaxial tensile strain promotes bone marrow-derived mesenchymal stem cells to differentiate into cardiomyocyte-like cells by mirna-27a: Int J Biochem Cell Biol, 2018; 99; 125-32
27. Zhang W, Kong C, Tong M, Maturation of human embryonic stem cell-derived cardiomyocytes (hesc-cms) in 3d collagen matrix: Effects of niche cell supplementation and mechanical stimulation: Acta Biomaterialia, 2017; 49; 204-17
28. Kroll K, Chabria M, Wang K, Electro-mechanical conditioning of human ipsc-derived cardiomyocytes for translational research: Prog Biophys Mol Biol, 2017; 130; 212-22
29. Shyu KG, Cellular and molecular effects of mechanical stretch on vascular cells and cardiac myocytes: Clin Sci (Lond), 2009; 116(5); 377-89
30. Kurazumi H, Kubo M, Ohshima M, The effects of mechanical stress on the growth, differentiation, and paracrine factor production of cardiac stem cells: PLoS One, 2011; 6(12); e28890
31. Ruan JL, Tulloch NL, Saiget M, Mechanical stress promotes maturation of human myocardium from pluripotent stem cell-derived progenitors: Stem Cells, 2015; 33(7); 2148-57
32. Qi Y, Li Z, Kong C-W, Uniaxial cyclic stretch stimulates trpv4 to induce realignment of human embryonic stem cell-derived cardiomyocytes: J Mol Cell Cardiol, 2015; 87; 65-73
33. Liaw NY, Zimmermann WH, Mechanical stimulation in the engineering of heart muscle: Adv Drug Deliv Rev, 2016; 96; 156-60
34. Shen N, Knopf A, Westendorf C, Steps toward maturation of embryonic stem cell-derived cardiomyocytes by defined physical signals: Stem Cell Rep, 2017; 9(1); 122-35
35. Lundy SD, Zhu W-Z, Regnier M, Structural and functional maturation of cardiomyocytes derived from human pluripotent stem cells: Stem Cells Dev, 2013; 22(14); 1991-2002
36. Ronaldson-Bouchard K, Ma SP, Yeager K, Advanced maturation of human cardiac tissue grown from pluripotent stem cells: Nature, 2018; 556(7700); 239-43
37. Tulloch NL, Muskheli V, Razumova MV, Growth of engineered human myocardium with mechanical loading and vascular coculture: Circ Res, 2011; 109(1); 47-59
38. Lu L, Mende M, Yang X, Design and validation of a bioreactor for simulating the cardiac niche: A system incorporating cyclic stretch, electrical stimulation, and constant perfusion: Tissue Eng Pt A, 2013; 19(3–4); 403-14
39. Lux M, Andrée B, Horvath T, In vitro maturation of large-scale cardiac patches based on a perfusable starter matrix by cyclic mechanical stimulation: Acta Biomaterialia, 2016; 30; 177-87
40. Abilez OJ, Tzatzalos E, Yang H, Passive stretch induces structural and functional maturation of engineered heart muscle as predicted by computational modeling: Stem Cells, 2018; 36(2); 265-77
41. Kensah G, Roa Lara A, Dahlmann J, Murine and human pluripotent stem cell-derived cardiac bodies form contractile myocardial tissue in vitro: Eur Heart J, 2013; 34(15); 1134-46
42. Reiser PJ, Portman MA, Ning X-H, Human cardiac myosin heavy chain isoforms in fetal and failing adult atria and ventricles: Am J Physiol Heart Circ Physiol, 2001; 280(4); H1814-20
43. Siedner S, Krüger M, Schroeter M, Developmental changes in contractility and sarcomeric proteins from the early embryonic to the adult stage in the mouse heart: J Physiol, 2003; 548(2); 493-505
44. Ng WA, Grupp IL, Subramaniam A, Cardiac myosin heavy chain mrna expression and myocardial function in the mouse heart: Circ Res, 1991; 68(6); 1742-50
45. Fishman GI, Hertzberg EL, Spray DC, Expression of connexin43 in the developing rat heart: Circ Res, 1991; 68(3); 782-87
46. Pervolaraki E, Dachtler J, Anderson RA, Ventricular myocardium development and the role of connexins in the human fetal heart: Sci Rep, 2017; 7(1); 12272
47. Eckardt D, Kirchhoff S, Kim JS, Cardiomyocyte-restricted deletion of connexin43 during mouse development: J Mol Cell Cardiol, 2006; 41(6); 963-71
48. Ya J, Erdtsieck-Ernste EBHW, Boer PAJd, Heart defects in connexin43-deficient mice: Circ Res, 1998; 82(3); 360-66
49. Ruan JL, Tulloch NL, Razumova MV, Mechanical stress conditioning and electrical stimulation promote contractility and force maturation of induced pluripotent stem cell-derived human cardiac tissue: Circulation, 2016; 134(20); 1557-67
50. Kensah G, Gruh I, Viering J, A novel miniaturized multimodal bioreactor for continuous in situ assessment of bioartificial cardiac tissue during stimulation and maturation: Tissue Eng Part C Methods, 2011; 17(4); 463-73
51. Massai D, Pisani G, Isu G, Bioreactor platform for biomimetic culture and in situ monitoring of the mechanical response of in vitro engineered models of cardiac tissue: Front Bioeng Biotechnol, 2020; 8; 733
52. Nguyen MD, Tinney JP, Ye F, Effects of physiologic mechanical stimulation on embryonic chick cardiomyocytes using a microfluidic cardiac cell culture model: Anal Chem, 2015; 87(4); 2107-13
53. Grandi E, Pandit SV, Voigt N, Human atrial action potential and ca2+ model: Sinus rhythm and chronic atrial fibrillation: Circ Res, 2011; 109(9); 1055-66
54. Yazawa M, Hsueh B, Jia X, Using induced pluripotent stem cells to investigate cardiac phenotypes in timothy syndrome: Nature, 2011; 471(7337); 230-34
55. Zhang XH, Haviland S, Wei H, Ca2+ signaling in human induced pluripotent stem cell-derived cardiomyocytes (ips-cm) from normal and catecholaminergic polymorphic ventricular tachycardia (cpvt)-afflicted subjects: Cell Calcium, 2013; 54(2); 57-70
56. Uzun AU, Mannhardt I, Breckwoldt K, Ca(2+)-currents in human induced pluripotent stem cell-derived cardiomyocytes effects of two different culture conditions: Front Pharmacol, 2016; 7; 300
57. Karakikes I, Ameen M, Termglinchan V, Human induced pluripotent stem cell-derived cardiomyocytes: Insights into molecular, cellular, and functional phenotypes: Circ Res, 2015; 117(1); 80-88
58. Gherghiceanu M, Barad L, Novak A, Cardiomyocytes derived from human embryonic and induced pluripotent stem cells: Comparative ultrastructure: J Cell Mol Med, 2011; 15(11); 2539-51
59. Rao C, Prodromakis T, Kolker L, The effect of microgrooved culture substrates on calcium cycling of cardiac myocytes derived from human induced pluripotent stem cells: Biomaterials, 2013; 34(10); 2399-411
60. Lee YK, Ng KM, Lai WH, Calcium homeostasis in human induced pluripotent stem cell-derived cardiomyocytes: Stem Cell Rev Rep, 2011; 7(4); 976-86
61. Hwang HS, Kryshtal DO, Feaster TK, Comparable calcium handling of human ipsc-derived cardiomyocytes generated by multiple laboratories: J Mol Cell Cardiol, 2015; 85; 79-88
62. White JPM, Cibelli M, Urban L, Trpv4: Molecular conductor of a diverse orchestra: Physiol Rev, 2016; 96(3); 911-73
63. Danciu TE, Adam RM, Naruse K, Calcium regulates the pi3k-akt pathway in stretched osteoblasts: FEBS Lett, 2003; 536(1–3); 193-197
64. Thodeti CK, Matthews B, Ravi A, Trpv4 channels mediate cyclic strain-induced endothelial cell reorientation through integrin-to-integrin signaling: Circ Res, 2009; 104(9); 1123-30
65. Barker N: The canonical wnt/β-catenin signalling pathway [M]//VINCAN E Wnt signaling: Pathway methods and mammalian models, 2008; 5-15, Totowa, NJ, Humana Press
66. Tzahor E, Wnt/β-catenin signaling and cardiogenesis: Timing does matter: Dev Cell, 2007; 13(1); 10-13
67. Renger A, Zafiriou M-P, Noack C, The four and a half lim-domain 2 controls early cardiac cell commitment and expansion via regulating beta-catenin-dependent transcription: Stem Cells, 2013; 31(5); 928-40
68. Banerjee I, Carrion K, Serrano R, Cyclic stretch of embryonic cardiomyocytes increases proliferation, growth, and expression while repressing tgf-β signaling: J Mol Cell Cardiol, 2015; 79; 133-44
69. Meza D, Abejar L, Rubenstein DA, A shearing-stretching device that can apply physiological fluid shear stress and cyclic stretch concurrently to endothelial cells: J Biomech Eng, 2016; 138(3); 4032550
70. Correia C, Koshkin A, Duarte P, 3D aggregate culture improves metabolic maturation of human pluripotent stem cell derived cardiomyocytes: Biotechnol Bioeng, 2018; 115(3); 630-44
71. Guo Y, Pu WT, Cardiomyocyte maturation: New phase in development: Circ Res, 2020; 126(8); 1086-106
In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387