03 June 2023: Review Articles
A Review of the Roles of Specialized Extracellular Vesicles, Migrasomes, and Exosomes in Normal Cell Physiology and Disease
Tingting ZhengABEFG, Shan XuCDE, Jinyuan XuCDEDOI: 10.12659/MSM.940118
Med Sci Monit 2023; 29:e940118






Abstract
ABSTRACT: Migrasomes are newly-discovered cellular organelle which are generated during cell migration and released from cells as extracellular vesicles (EVs), first described in 2015. Cellular contents are actively transported to migrasomes and released into extracellular space, then are taken up by other cells. Thus, migrasomes are proposed as a new mechanism for cell–cell communications, which show remarkable resemblance to exosomes, another classic EVs. The properties of exosomes in regulating intracellular communication have advanced their potential value in the therapeutic control of multiple diseases such as neurodegenerative conditions and cancer. Moreover, acting as potential biomarkers of various diseases, exosomes can be potentially valuable for diagnosis and assessment of the prognosis of patients with cancer or other diseases. Migrasomes are similar to exosomes in many characteristics. For instance, migrasomes can also mediate the lateral or horizontal transfer of materials among cells. On the other hand, although it is poorly understood, migrasomes show their own properties in normal cell physiology and disease. This review primarily summarizes recent advances in our understanding of the similarities and differences of migrasomes and exosomes in biogenesis, contents, and physiological and pathological effects on organisms, which may help us to have a better understanding of various types of EVs. This article aims to review of the roles of the specialized extracellular vesicles, migrasomes, and exosomes in normal cell physiology and disease.
Keywords: exosomes, Extracellular Vesicles, Cell Communication, Biomarkers, Humans, Neurodegenerative Diseases, Neoplasms
Background
Extracellular vesicles (EVs) are nanometer-ranged structures delimited by a lipid membrane, released by all cells under both normal and pathological conditions [1]. EVs are heterogeneous in origin, size, and cargo composition [1]. For instance, ectosomes are EVs that pinch off the surface of the plasma membrane via outward budding and are released directly from the plasma membrane [2]. Exosomes are another classic type of EVs, with an average diameter of ~100 nanometers with an endosomal origin [2]. It is formed by inward budding of the endosomal system and released after the multivesicular bodies (MVBs) fusion with the plasma membrane [3]. Exosomes are involved in a wide variety of physiological and pathological processes such as aging, immunity, and cancer [2]. Intercellular communication is an essential property of multicellular organisms. Exosomes also play important roles in communication among several organ systems [4] and can mediate intercellular communication by serving as vehicles for intercellular transfer of membrane and cytosolic proteins, lipids, and RNAs [5].
Recently, a new exosomelike organelle, named ‘migrasome’, has been attracting a great deal of attention [6]. The migrasomes, with diameters from 0.5 μm to 3 μm, contain numerous smaller vesicles with diameters of about 50–100 nm, which resemble opened pomegranates [6]. They derive from retraction fibers at the rear of migrating cells, and share some characteristics with EVs [6]. Migrasomes can also mediate the lateral or horizontal transfer of materials such as RNAs and proteins among cells [7]. Herein, we review the similarities and differences of migrasomes and exosomes in biogenesis, contents, intercellular communication, and as biomarkers for diseases, which may help us to have a better understanding of various types of EVs. This article aims to review of the roles of migrasomes and exosomes in normal cell physiology and disease.
Exosomes
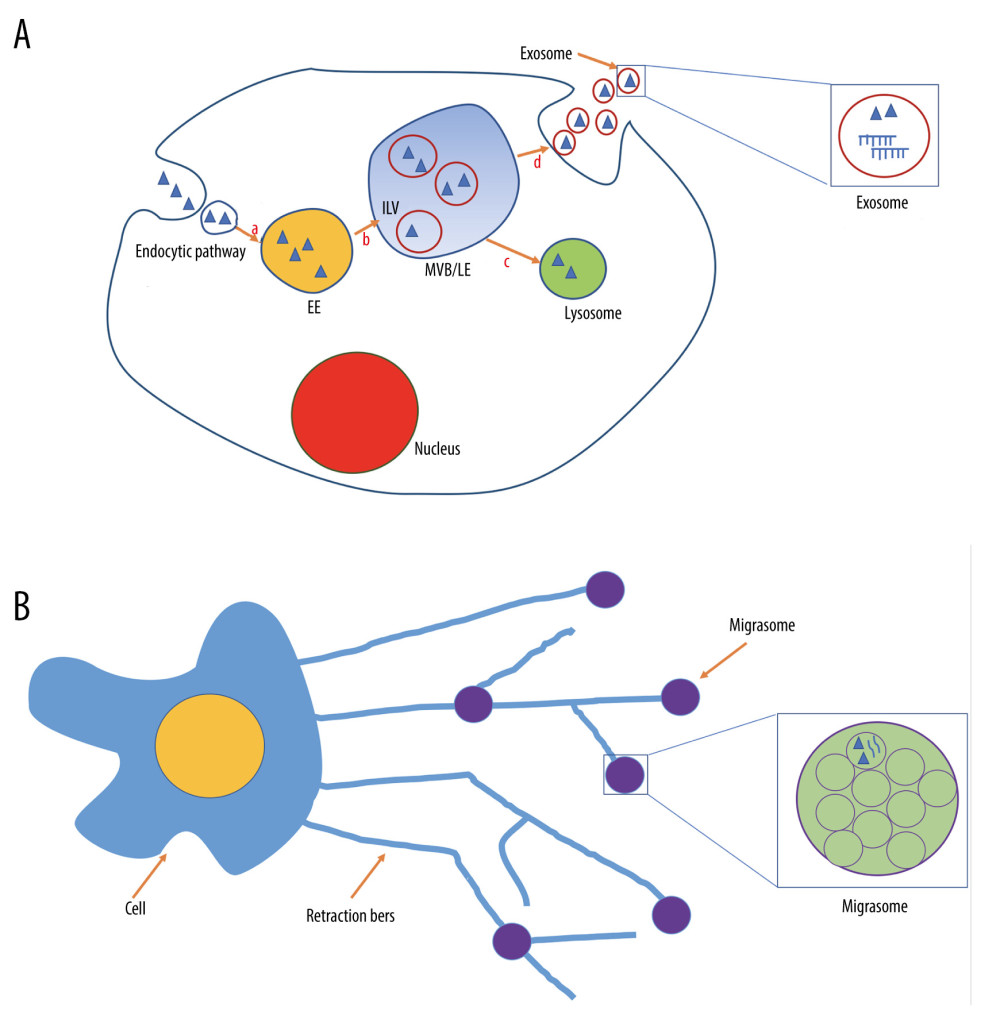
Exosomes are classic EVs typically 40–150 nm in diameter, generated in intracellular MVBs which contain intraluminal vesicles in the endosomal system [3]. The endosomal system is composed of primary endocytic vesicles, early endosomes (EEs), and late endosomes (LEs), which derive from EEs by stepwise maturation, and MVBs [8]. Upon formation, primary endocytic vesicles are transferred to EEs, which themselves mature into LEs. Then, the cargo can be targeted to MVBs, a type of LEs, which can fuse with plasma membrane to form exosomes [3,9,10] (Figure 1A). Exosomes can be secreted in vitro and in vivo by nearly all cell types and exist in almost all tissues, organs, and body fluids [11–14].
As a consequence of endosomal origin, nearly all exosomes contain proteins involved in endosome maturation (eg, Annexins, flotillin), MVBs biogenesis (eg, Alix and Tumor susceptibility gene 101 protein (TSG101)), heat-shock proteins, tetraspanins, and integrins [15]. Proteins of the tetraspanin family, characterized by containing 4 transmembrane domains, have 33 members [16]. Several members of the tetraspanin family have been identified in all kinds of exosomes independently of cell types, such as CD9, CD63, CD81, and CD82 [15], thus frequently serving as the surface markers of exosomes [17]. Exosomes also contain cell-type-specific proteins and lipids due to different cellular ancestries. For instance, exosomes derived from oligodendrocytes carry major myelin proteins [18]; the neural adhesion protein is only expressed in neural-derived exosomes [19].
Exosomes Mediate Intercellular Communication
Exosomes are important mediators of intercellular communication by delivering functional substances, including proteins, genetic cargo, RNAs, and lipids to recipient cells [2]. Exosomes are potential tools for cells to remove waste or toxic species such as prions, amyloid β, tau, and α-synuclein, which are involved in Creutzfeldt-Jacob disease, Alzheimer’s disease (AD), and Parkinson’s disease (PD) respectively [20–22]. Moreover, those toxic forms of aggregated proteins have been suggested to be spread from one cell to another through exosomes, causing an inflammatory cascade [20–22]. In addition, genetic cargo can be transferred between populations of tumor cells via exosomes, promoting drug resistance and progression of cancer [23–25]. The exosomes derived from THP1 cells carry miR-150 and deliver miR-150 into human microvascular endothelial cell lines (HMEC-1 cells). Then, these miR-150 significantly reduced c-Myb expression in HMEC-1 cells and promote cell migration [26].
Exosomes May be Used as Biomarkers
Exosomes may act as biomarkers for the diagnosis of various diseases. For instance, central nervous system-derived exosomal α-synuclein in plasma is substantially higher in PD patients, serving as a PD biomarker with high sensitivity and specificity [27]; exosome-derived biomarkers such as tau phosphorylated at r-181, cathepsin D, and Lysosome-associated membrane protein (LAMP-1) show high value in the diagnosis of AD and mild cognitive impairment [19,28]. Exosomes containing tumor-specific RNAs or proteins released from tumor cells can be potentially used for cancer diagnosis [29,30]. Exosomes from amniotic fluids may be used as a potent tool for prenatal diagnosis [31], and salivary exosomes may be useful in detecting Sjogren’s syndrome [32].
Migrasomes
Migrasomes are newly-discovered organelles produced by migrating cells [7]. Migracytosis is a new migration-dependent release mechanism for releasing cellular contents [6]. Cell migration is a complex process involving the coordinated action of signaling networks, cytoskeletal dynamics, cell adhesion, and membrane trafficking [33–35]. During migration, long tubular cytoplasmic extensions are pulled out from the rear end, which are called retraction fibers [36]. Then, membrane shedding can occur at the tips of, or at the intersections between, these thin retraction fibers, forming bulblike vesicles named “migrasomes” [6]. When retraction fibers break up, migrasomes are released from cells and exist in a package of unilamellar vesicles [6] (Figure 1B). Interestingly, migrasomes can release their internal vesicles to produce EV-like nanoparticles through self-rupture and via a process similar to cell plasma membrane budding [37]. Migrasome generation is strictly migration-dependent and is left in the tracks of migrating cells. Moreover, the number of migrasomes is related to cell migratory behavior [38].
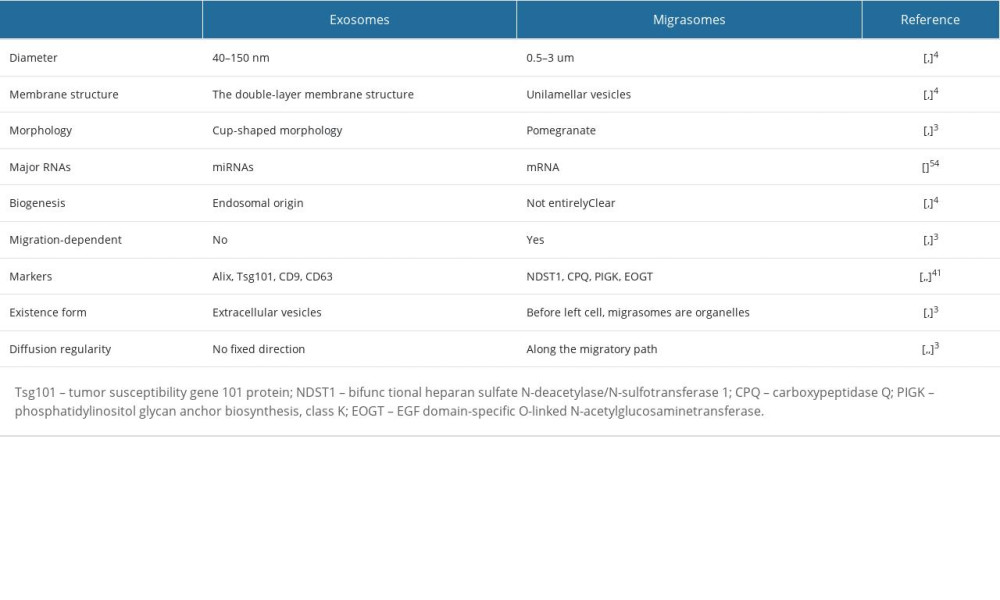
Similar to exosomes, migrasomes are present in multiple cell types, such as human and mouse cell lines, and are even found in macrophages, primary neurons, and embryonic stem cells [6,7,39]. In vivo, migrasome-like structures were found in various organs such as the lungs, eyes, and intestines [6], and even in postmortem brain tissue of stroke patients [40]. Migrasomes also been isolated from body fluid, such as serum and urine [41,42]. Before leaving the cell, migrasomes can carry out some cellular functions [6] (Table 1).
Compared to cell bodies, numerous proteins are enriched or depleted in migrasomes [43]. Similar to exosomes, migrasomal contents may be particularly associated with their biogenesis. The proteins enriched in migrasomes are involved in cell substrate adhesion, cell migration, lipid catabolic processes, protein glycosylation, and glycoprotein metabolic processes [41]. Moreover, as a migrating organelle, one feature of migrasomes is their enrichment in the contractile proteins actin and myosin, annexin proteins, and cytoskeleton [6,44].
The Contents of Migrasomes Compared to Exosomes
Different proteins, metabolites, and nucleic acids in migrasomes and exosomes may lead to a crucial difference in their physiological and pathological functions. There are similarities and differences of contents in migrasomes and exosomes. For instance, the tetraspanin family is enriched both in exosomes and migrasomes [45]. Huang et al observed that overexpression of 14 kinds of tetraspanins (including CD81 and CD82, which are specifically enriched in exosomes) enhanced migrasome formation in a dose-dependent manner [46]. However, tetraspanin CD9 and CD63, another 2 exosomal markers, showed no significant effect on migrasome formation [46]. Proteins of the tetraspanin family can segregate in the membrane plane into tetraspanin-enriched microdomains (TEMs) [47]. Cholesterol, another key component of TEMs, is also abundant on both migrasomes and exosomes [46,48]. Similar to tetraspanin, cholesterol is necessary for migrasome formation [46]. On the other hand, tetraspanin and cholesterol are sufficient for migrasome formation [46].
Integrins, a large family of transmembrane receptors, are composed of α and β subunits, and connect cells to the extracellular matrix (ECM) [49]. The integrin α5 and β1 are enriched in migrasomes, and can pair with their respective ECM partner to hold migrasomes on the ECM, emerging as an important principle for determining migrasomes formation and specific spatial and temporal distribution [39]. Although exosomes are always dissociated from cells and exist alone, they also contain integrins, including α5 and β1 [50]. For instance, exosomal integrin α5β1/asparaginyl endopeptidase complex, derived from epithelial ovarian cancer cells, plays an important role in mesothelial cell proliferation and migration, thus promoting peritoneal metastasis [50].
Interestingly, exosome release can affect the production of migrasomes. When Rab27a plasma membrane docking is inhibited, the migration of neural crest cells becomes slow and directionless [51]. The migration speed and persistence are positively correlated with migrasome formation by regulating retraction fibers [52].
Although there are many critical proteins in both migrasomes and exosomes, quantitative mass spectrometry suggests that the 2 structures share only 27% of their proteins [41]. In addition to tetraspanin 4 (TSPAN4) [53], Zhao et al identified 4 migrasome-specific potential biomarkers – NDST1 (bifunctional heparan sulfate N-deacetylase/N-sulfotransferase 1), CPQ (carboxypeptidase Q),PIGK (phosphatidylinositol glycan anchor biosynthesis, class K), and EOGT (EGF domain- specific O-linked N-acetylglucosaminetransferase) – which are abundant in migrasomes but are not present in exosomes [41]. On the other hand, exosomal markers such as Alix, Tsg101 and CD63 are found in small amounts in migrasomes [42,54,55] (Table 1).
In addition to proteins, RNAs are also present in migrasomes. In contrast to exosomal RNAs that mostly belong to small RNA species such as miRNAs, migrasomal RNAs are mainly composed of mRNA species that are long (>200 nt) [7,54] (Table 1). In addition, the expression profile of miRNAs in migrasomes also differs from that in exosomes. For instance, podocyte migrasomes contain more miR-1303, miR-548a, miR-490-5p, miR- 611, and miR-661 than podocyte exosomes, while exosomes have larger amounts of miR-221-3p, miR-144-3p, and miR-4286 than do migrasomes [42].
The Role of Migrasomes in Cell–Cell Communications
Cellular contents can be released through both exosomes and migrasomes. As described above, migrasomes can be released into the extracellular environment when retraction fibers break up [6]. Jiao et al suggested that damaged mitochondria can be transported into migrasomes and subsequently be cleared from migrating cells, thus maintaining the quality of the mitochondrial pool [56]. Similar to exosomes, intact migrasomes can be engulfed by surrounding cells, mediating release and transfer of cytoplasmic contents during cell migration [6]. Thus, migrasomes may be a newly-discovered mechanism for communication and transportation of cellular contents between cells [54]. Zhu et al found that full-length Pten mRNA can be transferred from migrasomes to recipient cells, then are translated into Pten proteins, which can modulate the pAKT level in recipient cells and inhibit the proliferation of recipient cells [54]. Migrasomal Pten protein itself can also be transferred into recipient cells, with functional consequence [54].
Normal Physiology of Migrasomes
Suppression of migrasome formation through knockdown of
Migrasomes in Diseases
Sodium chloride has been shown to induce marked migrasome formation of microglia/macrophages in an acute ischemic stroke model [40], and the extent of migrasome formation is correlated with the number of shrunk neurons [40]. Moreover, neurons are found in close vicinity to migrasomes, and double immunofluorescent staining revealed co-localization of microglia migrasomes and neuron fragments, suggesting a potential interaction between the migrasomes and surrounding neurons [40]. Migrasomes can be internalized by retinal pigmented epithelium (RPE), promoting RPE activation and proliferative vitreoretinopathy (PVR) progression by increasing the migration and proliferation ability of RPE [59]. TSPAN4 is abundant in the migrasome membrane and is one of the most effective tetraspanins for migrasome induction [46], stabilizing the migrasome structure and acting as a migrasome marker [6,60]. The expressionof migrasome-related genes such as
Migrasomes Act as Potential Biomarkers
Recent works by Liu et al discovered that podocytes can release migrasomes during migration [42]. Podocyte injury can trigger its motility [64,65], thus significantly promoting the release of migrasomes [42]. In vivo, higher levels of podocytes-derived migrasomes was detected in urine in a mouse nephropathy model than those in the control group, which was validated in human patients with kidney injury [42]. The increase of migrasomes occurs before the appearance of proteinuria [42]. Thus, levels of urinary migrasomes may serve as a potential diagnostic biomarker for early development of podocyte-based nephropathies.
Conclusions
The migrasomes and exosomes are similar in many aspects, and also have their own characteristics, and even show functional complementation. For instance, the originating cell source of exosomes in urine is complex, while urinary migrasomes are mainly derived from podocytes [42]. Therefore, it seems that urinary migrasome is more suitable as a noninvasive renal disease marker [42].
Future Directions
It is poorly understood how migrasomes travel so far, and much more research is needed to explore the physiological functions and pathological significance of migrasomes. For instance, the mechanisms underlying migrasome biogenesis and the pathways controlling migrasome formation are largely unclear. Moreover, it is unknown whether migrasomes carry disease-related proteins similar to exosomes, and if so, their role in various diseases. In addition, it is unclear whether migrasomes transfer toxic forms of aggregated proteins or genetic cargo between cells in diseases such as neurodegenerative disease and cancer. It is also unknown whether production of migrasomes from tumor cells is the major part of tumor metastasis. Exosomes may be engineered to deliver various therapeutic payloads, including chemotherapeutic agents, immune modulators, and short interfering RNAs, with an ability to direct their delivery to a desired target.
Much more research is needed to determine whether migrasomes have the potential to be engineered to deliver drugs, similar to exosomes.
References
1. Cocozza F, Grisard E, Martin-Jaular L, SnapShot: Extracellular vesicles: Cell, 2020; 182(1); 262-262e261
2. Kalluri R, LeBleu VS, The biology, function, and biomedical applications of exosomes: Science, 2020; 367(6478); eaauu6977
3. Dreyer F, Baur A, Biogenesis and functions of exosomes and extracellular vesicles: Methods Mol Biol, 2016; 1448; 201-16
4. Thery C, Zitvogel L, Amigorena S, Exosomes: Composition, biogenesis and function: Nat Rev Immunol, 2002; 2(8); 569-79
5. Maas SLN, Breakefield XO, Weaver AM, Extracellular vesicles: Unique intercellular delivery vehicles: Trends Cell Biol, 2017; 27(3); 172-88
6. Ma L, Li Y, Peng J, Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration: Cell Res, 2015; 25(1); 24-38
7. Yu S, Yu L, Migrasome biogenesis and functions: FEBS J, 2022; 289(22); 7246-54
8. Huotari J, Helenius A, Endosome maturation: EMBO J, 2011; 30(17); 3481-500
9. Heijnen HF, Schiel AE, Fijnheer R, Activated platelets release two types of membrane vesicles: Microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules: Blood, 1999; 94(11); 3791-99
10. Le Blanc I, Luyet PP, Pons V, Endosome-to-cytosol transport of viral nucleocapsids: Nat Cell Biol, 2005; 7(7); 653-64
11. Caby MP, Lankar D, Vincendeau-Scherrer C, Exosomal-like vesicles are present in human blood plasma: Int Immunol, 2005; 17(7); 879-87
12. Pisitkun T, Shen RF, Knepper MA, Identification and proteomic profiling of exosomes in human urine: Proc Natl Acad Sci USA, 2004; 101(36); 13368-73
13. Ogawa Y, Kanai-Azuma M, Akimoto Y, Exosome-like vesicles with dipeptidyl peptidase IV in human saliva: Biol Pharm Bull, 2008; 31(6); 1059-62
14. Vella LJ, Greenwood DL, Cappai R, Enrichment of prion protein in exosomes derived from ovine cerebral spinal fluid: Vet Immunol Immunopathol, 2008; 124(3–4); 385-93
15. Wubbolts R, Leckie RS, Veenhuizen PT, Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation: J Biol Chem, 2003; 278(13); 10963-72
16. Rubinstein E, The complexity of tetraspanins: Biochem Soc Trans, 2011; 39(2); 501-5
17. Meldolesi J, Exosomes and ectosomes in intercellular communication: Curr Biol, 2018; 28(8); R435-44
18. Kramer-Albers EM, Bretz N, Tenzer S, Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons?: Proteomics Clin Appl, 2007; 1(11); 1446-61
19. Goetzl EJ, Boxer A, Schwartz JB, Altered lysosomal proteins in neural-derived plasma exosomes in preclinical Alzheimer disease: Neurology, 2015; 85(1); 40-47
20. Yan M, Zheng T, Role of the endolysosomal pathway and exosome release in tau propagation: Neurochem Int, 2021; 145; 104988
21. Zheng T, Zhang Z, Activated microglia facilitate the transmission of alpha-synuclein in Parkinson’s disease: Neurochem Int, 2021; 148; 105094
22. Zheng T, Wu X, Wei X, The release and transmission of amyloid precursor protein via exosomes: Neurochem Int, 2018; 114; 18-25
23. Corcoran C, Rani S, O’Brien K, Docetaxel-resistance in prostate cancer: Evaluating associated phenotypic changes and potential for resistance transfer via exosomes: PLoS One, 2012; 7(12); e50999
24. Safaei R, Larson BJ, Cheng TC, Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells: Mol Cancer Ther, 2005; 4(10); 1595-604
25. O’Brien K, Rani S, Corcoran C, Exosomes from triple-negative breast cancer cells can transfer phenotypic traits representing their cells of origin to secondary cells: Eur J Cancer, 2013; 49(8); 1845-59
26. Zhang Y, Liu D, Chen X, Secreted monocytic miR-150 enhances targeted endothelial cell migration: Mol Cell, 2010; 39(1); 133-44
27. Shi M, Liu C, Cook TJ, Plasma exosomal alpha-synuclein is likely CNS-derived and increased in Parkinson’s disease: Acta Neuropathol, 2014; 128(5); 639-50
28. Xing W, Gao W, Lv X, The diagnostic value of exosome-derived biomarkers in Alzheimer’s disease and mild cognitive impairment: A meta-analysis: Front Aging Neurosci, 2021; 13; 637218
29. Properzi F, Logozzi M, Fais S, Exosomes: The future of biomarkers in medicine: Biomark Med, 2013; 7(5); 769-78
30. Lin J, Li J, Huang B, Exosomes: Novel biomarkers for clinical diagnosis: ScientificWorldJournal, 2015; 2015; 657086
31. Keller S, Rupp C, Stoeck A, CD24 is a marker of exosomes secreted into urine and amniotic fluid: Kidney Int, 2007; 72(9); 1095-102
32. Michael A, Bajracharya SD, Yuen PS, Exosomes from human saliva as a source of microRNA biomarkers: Oral Dis, 2010; 16(1); 34-38
33. Ridley AJ, Schwartz MA, Burridge K, Cell migration: integrating signals from front to back: Science, 2003; 302(5651); 1704-9
34. Ulrich F, Heisenberg CP, Trafficking and cell migration: Traffic, 2009; 10(7); 811-18
35. Friedl P, Wolf K, Plasticity of cell migration: A multiscale tuning model: J Cell Biol, 2010; 188(1); 11-19
36. Palecek SP, Schmidt CE, Lauffenburger DA, Horwitz AF, Integrin dynamics on the tail region of migrating fibroblasts: J Cell Sci, 1996; 109(Pt 5); 941-52
37. Ma Y, Li T, Zhao L, Isolation and characterization of extracellular vesicle-like nanoparticles derived from migrasomes: FEBS J, 2023 Online ahead of print
38. Lu P, Liu R, Lu D, Chemical screening identifies ROCK1 as a regulator of migrasome formation: Cell Discov, 2020; 6(1); 51
39. Wu D, Xu Y, Ding T, Pairing of integrins with ECM proteins determines migrasome formation: Cell Res, 2017; 27(11); 1397-400
40. Schmidt-Pogoda A, Strecker JK, Liebmann M, Dietary salt promotes ischemic brain injury and is associated with parenchymal migrasome formation: PLoS One, 2018; 13(12); e0209871
41. Zhao X, Lei Y, Zheng J, Identification of markers for migrasome detection: Cell Discov, 2019; 5; 27
42. Liu Y, Li S, Rong W, Podocyte-released migrasomes in urine serve as an indicator for early podocyte injury: Kidney Dis (Basel), 2020; 6(6); 422-33
43. Jiang D, Jiang Z, Lu D, Migrasomes provide regional cues for organ morphogenesis during zebrafish gastrulation: Nat Cell Biol, 2019; 21(8); 966-77
44. da Rocha-Azevedo B, Schmid SL, Migrasomes: A new organelle of migrating cells: Cell Res, 2015; 25(1); 1-2
45. Tavano S, Heisenberg CP, Migrasomes take center stage: Nat Cell Biol, 2019; 21(8); 918-20
46. Huang Y, Zucker B, Zhang S, Migrasome formation is mediated by assembly of micron-scale tetraspanin macrodomains: Nat Cell Biol, 2019; 21(8); 991-1002
47. Charrin S, le Naour F, Silvie O, Lateral organization of membrane proteins: Tetraspanins spin their web: Biochem J, 2009; 420(2); 133-54
48. Subra C, Laulagnier K, Perret B, Record M, Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies: Biochimie, 2007; 89(2); 205-12
49. Zaidel-Bar R, Job-splitting among integrins: Nat Cell Biol, 2013; 15(6); 575-77
50. Li X, Tang M, Zhu Q, The exosomal integrin alpha5beta1/AEP complex derived from epithelial ovarian cancer cells promotes peritoneal metastasis through regulating mesothelial cell proliferation and migration: Cell Oncol (Dordr), 2020; 43(2); 263-77
51. Gustafson CM, Roffers-Agarwal J, Gammill LS, Chick cranial neural crest cells release extracellular vesicles that are critical for their migration: J Cell Sci, 2022; 135(12); jcs260272
52. Fan C, Shi X, Zhao K, Cell migration orchestrates migrasome formation by shaping retraction fibers: J Cell Biol, 2022; 221(4); e202109168
53. Yu L, Migrasomes: The knowns, the known unknowns and the unknown unknowns: A personal perspective: Sci China Life Sci, 2021; 64(1); 162-66
54. Zhu M, Zou Q, Huang R, Lateral transfer of mRNA and protein by migrasomes modifies the recipient cells: Cell Res, 2021; 31(2); 237-40
55. Deniz IA, Karbanova J, Wobus M, Mesenchymal stromal cell-associated migrasomes: A new source of chemoattractant for cells of hematopoietic origin: Cell Commun Signal, 2023; 21(1); 36
56. Jiao H, Jiang D, Hu X, Mitocytosis, a migrasome-mediated mitochondrial quality-control process: Cell, 2021; 184(11); 2896-2910e2813
57. Mizoguchi T, Verkade H, Heath JK, Sdf1/Cxcr4 signaling controls the dorsal migration of endodermal cells during zebrafish gastrulation: Development, 2008; 135(15); 2521-29
58. Zhang C, Li T, Yin S, Monocytes deposit migrasomes to promote embryonic angiogenesis: Nat Cell Biol, 2022; 24(12); 1726-38
59. Wu L, Yang S, Li H, TSPAN4-positive migrasome derived from retinal pigmented epithelium cells contributes to the development of proliferative vitreoretinopathy: J Nanobiotechnology, 2022; 20(1); 519
60. Dharan R, Huang Y, Cheppali SK, Tetraspanin 4 stabilizes membrane swellings and facilitates their maturation into migrasomes: Nat Commun, 2023; 14(1); 1037
61. Qin Y, Yang J, Liang C, Pan-cancer analysis identifies migrasome-related genes as a potential immunotherapeutic target: A bulk omics research and single cell sequencing validation: Front Immunol, 2022; 13; 994828
62. Zheng Y, Lang Y, Qi B, Li T, TSPAN4 and migrasomes in atherosclerosis regression correlated to myocardial infarction and pan-cancer progression: Cell Adh Migr, 2023; 17(1); 14-19
63. Zheng Y, Lang Y, Qi B, TSPAN4 is a prognostic and immune target in Glioblastoma multiforme: Front Mol Biosci, 2022; 9; 1030057
64. Ivanova EA, Arcolino FO, Elmonem MA, Cystinosin deficiency causes podocyte damage and loss associated with increased cell motility: Kidney Int, 2016; 89(5); 1037-48
65. Cechova S, Dong F, Chan F, MYH9 E1841K mutation augments proteinuria and podocyte injury and migration: J Am Soc Nephrol, 2018; 29(1); 155-67
In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387