01 September 2020: Animal Study
Chlorogenic Acid Alleviates Allergic Inflammatory Responses Through Regulating Th1/Th2 Balance in Ovalbumin-Induced Allergic Rhinitis Mice
Feilin Dong1ABE, Jun Tan1CDE, Yi Zheng1DF*DOI: 10.12659/MSM.923358
Med Sci Monit 2020; 26:e923358






Abstract
BACKGROUND: Allergic rhinitis (AR) is a prevalent atopic disorder caused by immune imbalance. Chlorogenic acid (CGA) has antibacterial, antiviral, antioxidative and immunoregulatory effects, but its role in anaphylactic disease remains unclear. The current study aimed to investigate the function of CGA in AR.
MATERIAL AND METHODS: AR mice models were induced with ovalbumin (OVA) by orally administrating the mice with 50 mg/kg (L-CGA), 100 mg/kg (M-CGA), or 200 mg/kg (H-CGA) of CGA. The number of nasal rubbings and sneezes was recorded. Afterward, the mice were sacrificed for the collection of blood, nasal lavage fluid (NALF), and nasal tissues. The cells in NALF were counted by hemocytometer and stained by Diff-Quick. Nasal mucosa was observed by H&E staining. ELISA testing was conducted for detecting the levels of anti-OVA antibodies and Th1/Th2-related cytokine. Quantitative real-time polymerase chain reaction experiments were conducted to determine mRNA expressions of Th1/Th2-related cytokines.
RESULTS: In the OVA-induced AR mice, CGA treatment reduced nasal rubbing and sneezing, and also suppressed the number of total cells, eosinophils, neutrophils, lymphocytes, macrophages, and epithelial cells in NALF. OVA-induced up-regulation of nasal mucosa thickness was inhibited by CGA, and the effects of OVA on IgE, IgG1, and IgG2a were reversed by CGA. The regulatory effects of OVA on mRNA expressions and levels of Th1/Th2-related cytokines were abolished by CGA treatment in AR mice.
CONCLUSIONS: CGA can alleviate allergic inflammatory responses through regulating Th1/Th2 balance in OVA-induced allergic rhinitis mice.
Keywords: Cytokines, Rhinitis, Allergic, Seasonal, Systemic Inflammatory Response Syndrome, Chlorogenic Acid, Enzyme-Linked Immunosorbent Assay, Eosinophils, Inflammation Mediators, Nasal Mucosa, Ovalbumin, Real-Time Polymerase Chain Reaction, Rhinitis, Allergic, Th1-Th2 Balance
Background
Rhinitis is a disorder defined as nasal mucosal inflammation, and allergic rhinitis (AR) is the most common type, which could lead to the increase of type 2 helper (Th2) cells and the decrease of type 1 helper (Th1) cells [1]. AR affects 10% to 20% of populations of all ages worldwide [2–4]. Patients suffering from rhinitis may show symptoms of AR, such as poor sleeping quality and performance at work or school, and numerous comorbidities such as headaches, allergic conjunctivitis, otitis media, and asthma [5]. However, the current management of AR as a chronic condition is far from satisfactory [6]. Moreover, AR patients treated by various medications such as antihistamines, antileukotriene, and intranasal corticosteroids are still experiencing treatment failures or dissatisfactory results [7]. Thus, developing effective and novel treatment methods is vitally important.
Normal cellular and humoral immune responses are regulated by CD4+ T helper cells in reaction to various pathogens in the progression of many diseases [8]. Th1 and Th2 cells are subsets of CD4+ T helper cells, and the balance between them plays a vital role in allergic and autoimmune disorders via mediating the secretion of Th1-related cytokines interferon (IFN)-γ, interleukin (IL)-2, tumor necrosis factor (TNF)-β, and Th2-related cytokines [9,10]. Imbalance in the CD4+ T cell subset and the accumulation of eosinophils and mast cells in the nasal mucosa is characteristic of AR [11]. Therefore, maintaining the balance of Th1 and Th2 cells and inhibiting the accumulation of eosinophil and mast cells in the nasal mucosa are the keys to the treatment of this disease.
Chlorogenic acid (CGA) is a caffeoylquinic acid (CQA) isomer which is known as 5-CQA, according to the guidelines of the International Union of Pure and Applied Chemistry (IUPAC). The chemical structure of CGA is demonstrated in Figure 1A. CGA can be naturally found in tea and the extraction of green coffee [12]. Researches showed that CGA not only has neuro-protective and vascular-protective effects [13,14], but also has anti-inflammatory, antiviral, antilipidemic, and antioxidative functions [15–18]. It has been reported that dietary CGA plays an anti-inflammatory role in inhibiting IL-8 production induced by H2O2 and TNF-α in Caco-2 cells [18]. In addition, CGA can act as a potent anti-arthritic agent by suppressing T cells and Th1-related cytokines, thus enhancing the immune system via increasing IL-4 and IL-10 [19]. It has been reported that CGA potentiated IgE production
To the best of our knowledge, the current study is the first to explore the function of CGA in an OVA-induced AR mice model involving Th1/Th2 balance, and we aimed to develop a novel treatment for the management of AR.
Material and methods
ANIMALS:
A total of 60 male BALB/c mice (catalog ID: BALB-M; male, 6-week-old) were purchased from Taconic Biosciences (NY, USA). The mice were housed in a stable environment (temperature: 23±3°C; relative humidity: 50±10%; 12-h light/dark cycles) for 1 week before the experiments. All the animal experiments were performed in accordance with the guidelines of the China Council on Animal Care and Use. This study was approved by the Committee of Experimental Animals. All possible efforts were made to minimize the pain and discomfort of the animals. The animal experiments were performed in the Zhejiang Provincial People’s Hospital.
PREPARATION OF CGA:
CGA (catalog ID: A10202) used for experimentation was obtained from Adooq Bioscience (CA, USA) with purity higher than 98%. CGA powder was dissolved in saline and prepared to concentrations of 50 mg/kg (low (L)-CGA group), 100 mg/kg (medium (M)-CGA group), and 200 mg/kg (high (H)-CGA group). All the reagents were freshly prepared before administration of daily treatments. Ten mg/kg cromolyn disodium salt hydrate (DSGC) was used in a positive control group.
ESTABLISHMENT AND GROUPING OF THE AR MOUSE MODEL:
Mice were divided into 6 groups (n=10 per group): control, OVA, DSGC, L-CGA, M-CGA, and H-CGA. Groups are described in Figure 1B. OVA group: mice were sensitized with 50 μg OVA (S7951, Sigma-Aldrich, USA) and 1 mg aluminum hydroxide (239186, Sigma-Aldrich, USA) on days 0, 7, and 14, and 1 week after the last sensitization; namely, mice received an intranasal challenge with 10% OVA on days 21 to 27. Mice were treated 30 min before the intranasal challenge of OVA, and from days 15 to 27, mice were given saline once daily. DSGC group: the sensitization process in the mice was the same as in the OVA group, then from days 15 to 27, mice were orally administered 10 mg/kg DSGC once daily. L-CGA, M-CGA and H-CGA groups: the sensitization process in the mice was the same as in the OVA group, then from days 15 to 27, mice were orally administered 50, 100, and 200 mg/kg CGA once daily, respectively. control group: the mice were given an equal volume of distilled water instead of the drug. Moreover, mice in all groups received intranasal challenges with 10% OVA once a day 30 minutes (min) after the treatment, and the number of times of nasal rubbing and sneezing were recorded 15 min after the challenge. Finally, the mice were sacrificed on day 26 and examined on day 27.
NASAL LAVAGE FLUID COLLECTION AND CELL COUNTING:
Blood was harvested via cardiac puncture from the sacrificed mice, and serum was collected by centrifugation. The trachea was partially resected, and then an 18-gauge catheter was inserted into the trachea towards upper airway into the nasopharynx. Next, the nasal passage was perfused with 1 mL pre-cooled phosphate buffer saline (PBS, Gibco, California, US). Then, nasal lavage fluid (NALF) was collected and centrifuged to obtain the supernatants for cytokines measurements. The cells were suspended in PBS and the total number of cells was counted with a hemocytometer (Solarbio, Beijing, China). An amount of 150 μL NALF was centrifuged onto glass slides using a Cytopro Cytocentrifuge Series 2 cytospin device (ELITech Group, Puteaux, France) at 1000 rmp for 10 min at 4°C. A Diff-Quick stain kit (Solarbio, Beijing, China) was used for cell staining in accordance with the manufacturer’s protocol. Finally, a Nicolet Continuum Infrared Microscope (Thermo Scientific, California, USA) was used for observation.
HISTOLOGICAL EXAMINATION:
The heads of the mice were fixed in neutral 10% formalin fixative (PH7.4, Solarbio, Beijing, China), successively dehydrated by 80%, 90%, 95% and 100% ethyl alcohol (Sigma, California, US), and made transparent by xylenes I & II (Sigma, CA, USA). Next, the tissues were paraffin-embedded (Chemical Book, Beijing, China), and cut into 5-μm-thick sections, washed with xylenes and ethyl alcohol for debenzolization and rehydration, and then stained with a hematoxylin and eosin (H&E) staining kit (ab245880, Abcam, USA). Next, dehydration and transparentizing of the tissue slices were conducted again using ethyl alcohol and xylenes. Finally, the samples were sealed in neutral balsam (Macklin, Shanghai, China). The epithelial damages were observed under a microscope with 1000× magnification.
MEASUREMENT OF CYTOKINE LEVELS:
NALF was centrifugalized to collect the supernatant. The blood was collected from the vena cava, and then serum was collected via centrifugation. Nasal tissues of sacrificed mice were stored at −70°C, and then homogenated and centrifuged to collect the supernatant. The anti-OVA Ig (immunoglobulin) E, anti-OVA IgG1 and anti-OVA IgG1 levels in serum, and IFN-γ, IL-12, IL-4, IL-5, and IL-13 levels in the NALF and nasal mucosa were measured through enzyme-linked immunosorbent assays (ELISA) using an anti-ovalbumin IgE (mouse) ELISA kit (500840, Cayman, MI, USA), anti-ovalbumin IgG1 (mouse) ELISA kit (500830, Cayman, MI, USA), mouse anti-ovalbumin (Gal d 2) IgG2a ELISA Kit (600-120-02A, Biolead, China), mouse IFN-γ ELISA kit (SEKM-0031, Solarbio, China), mouse IL-12p40 ELISA kit (SEKM-0012, Solarbio, China), mouse IL-13 ELISA kit (SEKM-0014, Solarbio, China), mouse IL-5 ELISA kit (SEKM-0006, Solarbio, China), and mouse IL-4 ELISA kit (SEKM-0005, Solarbio, China) according to the manufacturers’ instructions.
QUANTITATIVE REAL-TIME POLYMERASE CHAIN REACTION:
Total mRNAs were isolated from the NALFs and nasal mucosa by using TRIzol Reagent (Invitrogen, CA, USA). Then the first strand cDNA was synthesized using a high-capacity RNA-to-cDNA kit (Applied Biosystems, CA, USA). Quantitative real-time polymerase chain reaction (qPCR) was performed using 20 μL SYBR Select Master Mix (Applied Biosystems, CA, USA) in a 7500 Fast Real-Time PCR System, laptop (Applied Biosystems, CA, USA) as follows: at 95°C for 2 min, followed by 40 cycles at 95°C for 3 sec, and at 60°C for 30 sec. The relative expression of each mRNA was calculated by comparative cycle threshold (CT) method (2−ΔΔCT) [23]. The primers used are listed in Table 1.
STATISTICAL ANALYSIS:
The data are presented as mean±standard error and were analyzed in SPSS, v22.0 (IBM, USA). Statistical differences among the experimental groups were compared by one-way analysis of variance followed by Bonferroni’s post hoc test.
Results
OVA-INDUCED AR SYMPTOMS IN THE MICE WERE INHIBITED BY CGA:
Figure 1A shows the chemical structure of CGA. Figure 1B shows the simplified flowchart of the treatments. The effects of CGA on the early-phase allergic symptoms were detected by counting the number of nasal rubbings and sneezes 15 min after the OVA intranasal challenge on day 25. As shown in Figure 1C and 1D, the number of nasal rubbings and sneezes in the OVA group were significantly increased compared with those in the control group (P<0.001). However, allergic nasal symptoms were greatly alleviated in CGA-administrated group compared with those in the OVA group. DSGC also had an inhibitory effect on AR symptoms induced by OVA.
INFILTRATION OF DIFFERENTIAL INFLAMMATORY CELLS IN OVA-INDUCED MICE WAS REDUCED BY CGA IN NALF AND IN EPITHELIAL CELL DISRUPTION:
Diff-Quick staining results showed that loss of eosinophils (red arrow) and epithelial cells from inflammatory tissues (black arrow) were remarkably increased by OVA, but were greatly reversed by DSGC and CGA treatment in AR mice (Figure 2A). Moreover, the number of total cells and different inflammatory cells in the NALF were significantly increased in the OVA group. Nevertheless, oral administration of CGA (50, 100, and 200 mg/kg) and DSGC (10 mg/kg) effectively reduced these inflammatory cells (P<0.001; Figure 2B–2F). Thus, these results indicated that CGA improved the allergic inflammatory responses of AR in mice.
CGA HAD PROTECTIVE EFFECTS ON NASAL MUCOSA IN OVA-INDUCED AR MICE:
The general structure of nasal mucosa was observed through H&E staining (P<0.001; Figure 3A), and we found that subepithelial inflammatory cells in the nasal septum were increasingly accumulated; moreover, the nasal mucosa was aberrantly thickened in the OVA group compared with that of the control group. However, the nasal mucosa layers in AR mice treated by CGA and DSGC were relatively much thinner than those in the OVA group, suggesting that CGA might protect AR mice from nasal morphological abnormalities.
THE ANTI-OVA SPECIFIC ANTIBODIES IN SERUM WAS REGULATED BY CGA:
The anti-OVA specific antibodies in serum were assayed by ELISA to determine the function of CGA in allergic inflammatory responses (P<0.05; Figure 3B–3D). The experimental results demonstrated that anti-OVA IgE and anti-OVA IgG1 levels were markedly increased by OVA, whereas the level of anti-OVA IgG2a was down-regulated by OVA compared with that of the control group. However, CGA treatment decreased the anti-OVA IgE and anti-OVA IgG1 levels, but increased the level of anti-OVA IgG2a in AR mice serum compared with those measured in the OVA group. Moreover, the regulatory effects of DSGC were similar to CGA in the experiments.
CGA REGULATED THE BALANCE OF TH1/TH2-RELATED CYTOKINES IN NALF AND NASAL MUCOSA:
The levels of IFN-γ, IL-12, IL-4, IL-5, and IL-13 cytokines in the NALF and nasal tissues were measured by conducting ELISA (P<0.05; Figure 4A–4J) and qRT-PCR (P<0.05; Figure 5A, 5B) to study the modulatory functions of CGA. In the NALF and nasal tissues of the mice, OVA greatly promoted IL-4, IL-5, and IL-13 levels but inhibited IFN-γ and IL-12 levels compared with those of the control group. However, the levels of IL-4, IL-5, and IL-13 in the NALF and nasal tissues were significantly down-regulated in the L-CGA, M-CGA, and H-CGA groups. In addition, IFN-γ levels were increased in both the NALF and nasal tissues with the treatment of 50, 100, and 200 mg/kg CGA, while the levels of IL-12 were increased in the NALF with the treatment of 50, 100, and 200 mg/kg CGA, whereas IL-12 increased in nasal tissues with 200 mg/kg CGA only. The regulatory function of DSGC was similar to that of CGA. Noticeably, all the effects of OVA on Th1/Th2-related cytokines in the NALF and nasal tissues were reversed by CGA. Furthermore, OVA-induced increases of mRNA expressions of IL-4, IL-5, and IL-13, as well as OVA-induced decreases of mRNA expressions of IFN-γ and IL-12, were abolished by DSGC and CGA in NALF and nasal tissues; however, low dose of CGA (50 mg/kg) had no significant effect on mRNA expressions of IL-5 in nasal tissues.
Discussion
As one of the remarkable components of traditional Chinese medicines, CGA is well known for its diverse antioxidative, antimicrobial, anti-inflammatory, and hepato-protective pharmacological activities, but the anti-allergic function of CGA in AR treatment is rarely studied. In this study, an OVA-induced mice model was designed to study this potential function of CGA. Nasal symptoms, nasal mucosa thickness, OVA-specific antibodies, Th1/Th2-regulated cytokines, and inflammatory cells were detected to explore the effects of CGA on allergic inflammatory responses in AR.
AR causes nasal mucosal inflammation combined with tissue influx of eosinophils and basophils [2]. In the early stage of AR inflammation, histamine secreted by mast cells induces symptoms such as nasal itching and sneezing [24,25]. In the current study, our results showed that oral administration of CGA effectively alleviated the nasal symptoms in the OVA-induced mice model, and the effects of high-dose CGA in relieving nasal symptoms of OVA-induced mice were more obvious, suggesting that CGA might have anti-inflammatory effects on the early phase of AR.
Moreover, Th2-mediated acquired type 2 immune responses are induced by allergen and allergen-specific IgE antibodies. The antigen-specific effector is regulated by IgE via binding to the high-affinity receptor for IgE, which is expressed on the surface of mast cells and basophilic granulocytes [26]. A previous study showed that an eosinophil count of >0.3 per high-power field in nasal smears is a highly specific criterion for AR diagnosis [27], thus, the effect of CGA can actually be reflected through the IgE level and mast cells numbers in early-phase reactions of AR. In our study, the oral administration of CGA significantly reduced total cells and inflammation-related cells, including eosinophils, neutrophils, lymphocytes, and macrophages in NALF, in a dose-independent manner as compared with those of the OVA group. Moreover, the OVA-induced increases of anti-OVA IgE and anti-OVA IgG1 were largely reversed by CGA treatment, while the previously down-regulated anti-OVA IgG2a induced by OVA was up-regulated by CGA. Thus, all of these results indicate that oral administration of CGA may play a protective role in AR treatment via inhibiting allergic inflammatory responses with the regulation of OVA-specific antibodies and mast cells.
After exposure to an allergen for 2 to 9 h, late-phase reactions were generally developed, and the chronic allergic inflammation occurred with repeated allergen exposure. At this stage, the numbers of eosinophils, neutrophils, lymphocytes, and macrophages were increased, and then inflammatory cells infiltrated through the cavum nasi. Further, mediators derived from eosinophils will cause epithelial damage and nasal mucosal swelling [24,25,28]. In our study, the results from Diff-Quick staining experiments demonstrated that the loss of eosinophils and epithelial cells from inflammatory nasal tissues was significantly increased in the OVA group. However, CGA administration greatly reduced the cell numbers in the AR mice model. Moreover, it could be observed that the OVA-induced AR mice had abnormally thicker nasal mucosa than the mice in the control group; however, the aberrancy was reversed by CGA oral treatment, and the effect of high-dose CGA was most obvious. Thus, these results elucidated that CGA may prevent inflammatory cells from accumulation and inhibit eosinophil-derived regulators from activation, so that the nasal mucosa could be protected against hyperplasia in AR mice induced by OVA.
AR is an inflammatory disease caused by specific IgE-manipulated reactions in response to inhaled allergens, which are driven by Th2 cells [2]. Thus, Th2 cells play a pivotal role in the response to allergic inflammation through secreting Th2-related cytokines and promoting IgE-producing B-cells. IgE released by B-cells combines with the surface of mast cells, which further leads to the release of histamines [29,30]. IL-4, IL-5, and IL-13 are Th2-related cytokines that have been shown to regulate eosinophil growth and differentiation. Moreover, IL-13 can also promote inflammation, subepithelial fibrosis, and eotaxin production [31]. In the present study, CGA treatment greatly inhibited the levels of IL-4, IL-5, and IL-13 in the NALF and nasal mucosa, which may have resulted in the further suppression of activation of inflammatory cells. Similarly, the mRNA expressions of IL-4, IL-5, and IL-13 were also reduced by oral administration of CGA to OVA-induced mice. Additionally, Th1-associated cytokines IFN-γ and IL-12 have been reported to activate the Th1 differentiation and suppress the development of Th2 [32,33]. Our data showed that the levels and mRNA expressions of IFN-γ and IL-12 were inhibited by OVA compared with those of the control group, but were promoted after oral administration of CGA to the AR mice, suggesting that CGA might be effective in regulating the allergic inflammatory responses of AR through the mediation of Th1/Th2 balance.
In this study, OVA-induced mice were treated with different doses of CGA, and it was found that all concentrations of CGA could inhibit nasal mucosal thickness and regulate Th1/Th2 balance to a certain extent to reduce allergic inflammatory response, and low, medium, and high doses of CGA showed an enhanced trend in alleviating the effect of OVA-induced AR. This suggests that the use of 200 mg/kg CGA in the treatment of AR may be most significant. In addition, DSGC was used as the positive control, and the effect of CGA treatment on regulating nasal symptoms, inflammatory cell infiltration (eosinophils, neutrophils, and macrophages), nasal mucosal thickness, specific antibodies, and Th1/th2-related cytokine levels in OVA-induced AR mice was stronger than that of DSGC treatment. Thus, CGA treatment has better clinical significance in improving the allergic inflammatory response of AR mice induced by OVA. However, this study also has some limitations. It remains to be seen whether higher concentrations of CGA treatment have a better effect or a significant toxic effect. Whether CGA can reduce allergic inflammation in AR mice by regulating a signaling pathway also remains to be investigated.
Conclusions
In conclusion, our study demonstrated that CGA alleviated allergic inflammatory responses through the suppression of inflammatory cells, protected nasal tissues from hyperplasia, and prevented epithelial cells from inflammatory infiltration through the mediation of Th1/Th2 balance in OVA-induced AR mice. Therefore, CGA is a potential candidate for AR treatment.
Figures
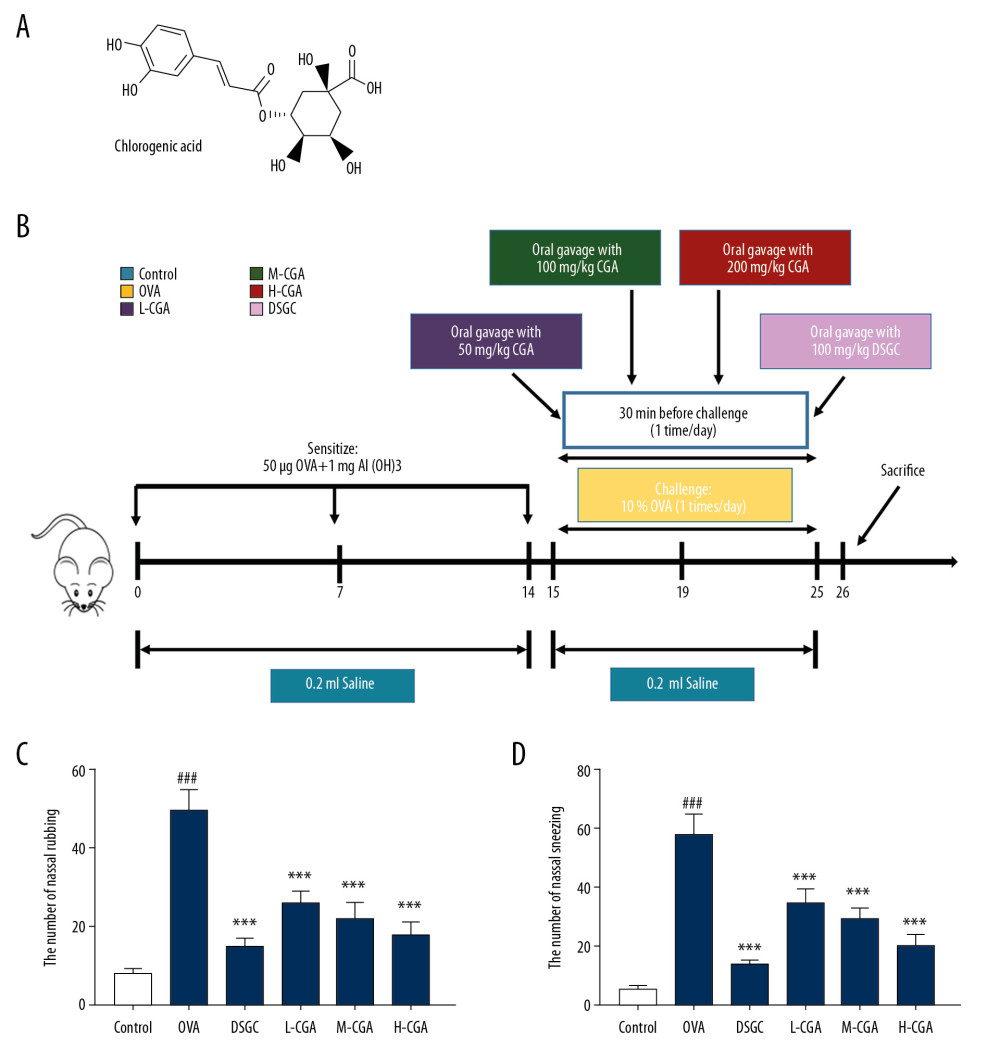 Figure 1. Chlorogenic acid (CGA) relieved the symptom of allergic rhinitis (AR) induced by ovalbumin (OVA) in mice. (A) The chemical formula of CGA is shown in the figure. (B) The mice were sensitized on days 0, 7, and 14 by OVA and Al(OH)3, and then orally gavaged from day 15 to 25 (11 days) with CAG (50, 100, and 200 mg/kg), Cromolyn disodium salt hydrate (DSGC, 100 mg/kg), or isopycnic distilled water once a day. Next, the mice were challenged with saline containing 10% OVA via nose drops 30 minutes (min) after gavage. Before sacrifice on day 26, (C, D), nasal-rubbing and -sneezing frequencies of the mice in different groups were counted on day 25. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA.
Figure 1. Chlorogenic acid (CGA) relieved the symptom of allergic rhinitis (AR) induced by ovalbumin (OVA) in mice. (A) The chemical formula of CGA is shown in the figure. (B) The mice were sensitized on days 0, 7, and 14 by OVA and Al(OH)3, and then orally gavaged from day 15 to 25 (11 days) with CAG (50, 100, and 200 mg/kg), Cromolyn disodium salt hydrate (DSGC, 100 mg/kg), or isopycnic distilled water once a day. Next, the mice were challenged with saline containing 10% OVA via nose drops 30 minutes (min) after gavage. Before sacrifice on day 26, (C, D), nasal-rubbing and -sneezing frequencies of the mice in different groups were counted on day 25. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA. 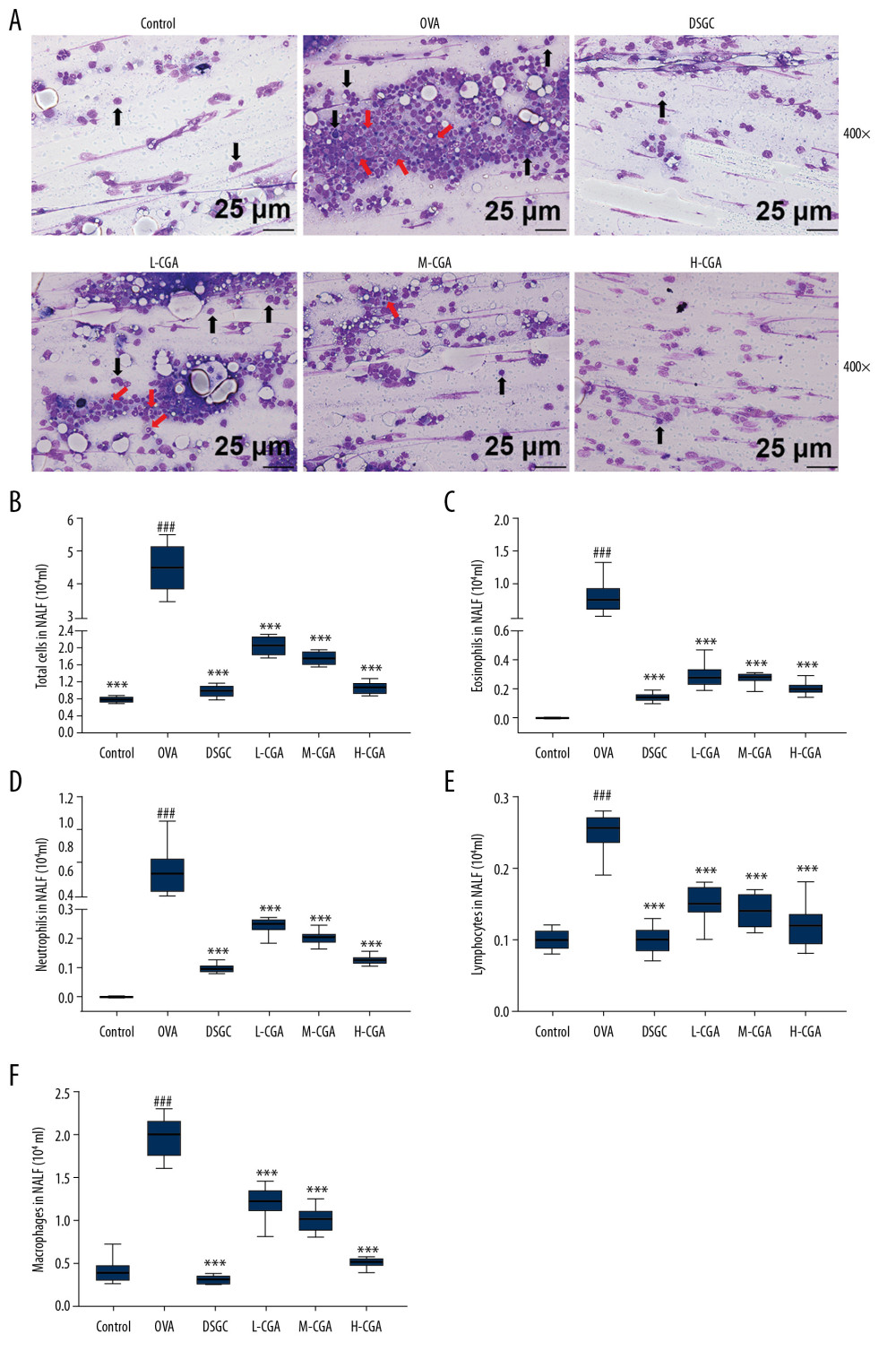 Figure 2. Chlorogenic acid (CGA) reduced the infiltration of differential inflammatory cells in nasal lavage fluid (NALF) and prevented epithelial cells disruption. NALF was collected immediately after sacrifice, and the cells were isolated by cytospin. (A) Cells from different groups were stained by Diff-Quick. Red arrows indicate eosinophils. Black arrows indicate epithelial cells lost from inflammatory tissues. (B) The number of total cells in NALF. (C) The number of eosinophils in NALF. (D) The number of neutrophils in NALF. (E) The number of lymphocytes in NALF. (F) The number of macrophages in NALF. Scale bar: 25μm, ×400 magnification. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA.
Figure 2. Chlorogenic acid (CGA) reduced the infiltration of differential inflammatory cells in nasal lavage fluid (NALF) and prevented epithelial cells disruption. NALF was collected immediately after sacrifice, and the cells were isolated by cytospin. (A) Cells from different groups were stained by Diff-Quick. Red arrows indicate eosinophils. Black arrows indicate epithelial cells lost from inflammatory tissues. (B) The number of total cells in NALF. (C) The number of eosinophils in NALF. (D) The number of neutrophils in NALF. (E) The number of lymphocytes in NALF. (F) The number of macrophages in NALF. Scale bar: 25μm, ×400 magnification. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA. 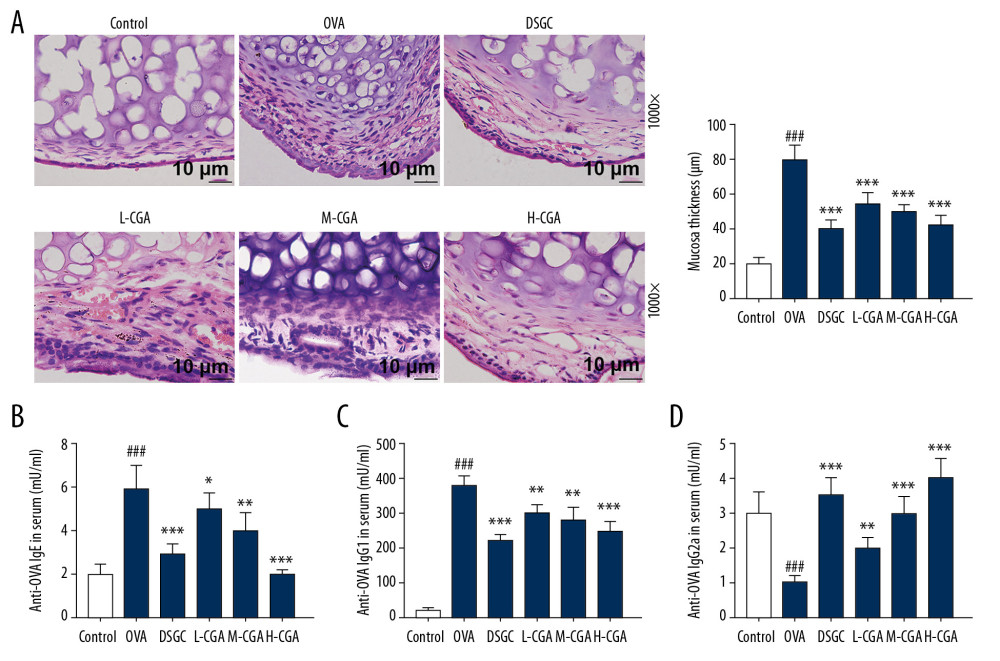 Figure 3. Chlorogenic acid (CGA) reduced mucosa thickness and mucus secretions in the nasal mucosa of mice. (A) Histological features of the nasal mucosa were identified by hematoxylin-eosin (HE) staining, and the mucosa thickness of nasal septum was measured. Enzyme-linked immunosorbent assays (ELISA) were performed to detect the levels of anti-OVA specific IgE (B), anti-OVA IgG1 (C) and anti-OVA IgG2a (D) in the serum of the mice. Scale bar: 10μm, ×1000 magnification. n=10 each group. ### P<0.001 vs. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA.
Figure 3. Chlorogenic acid (CGA) reduced mucosa thickness and mucus secretions in the nasal mucosa of mice. (A) Histological features of the nasal mucosa were identified by hematoxylin-eosin (HE) staining, and the mucosa thickness of nasal septum was measured. Enzyme-linked immunosorbent assays (ELISA) were performed to detect the levels of anti-OVA specific IgE (B), anti-OVA IgG1 (C) and anti-OVA IgG2a (D) in the serum of the mice. Scale bar: 10μm, ×1000 magnification. n=10 each group. ### P<0.001 vs. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA. 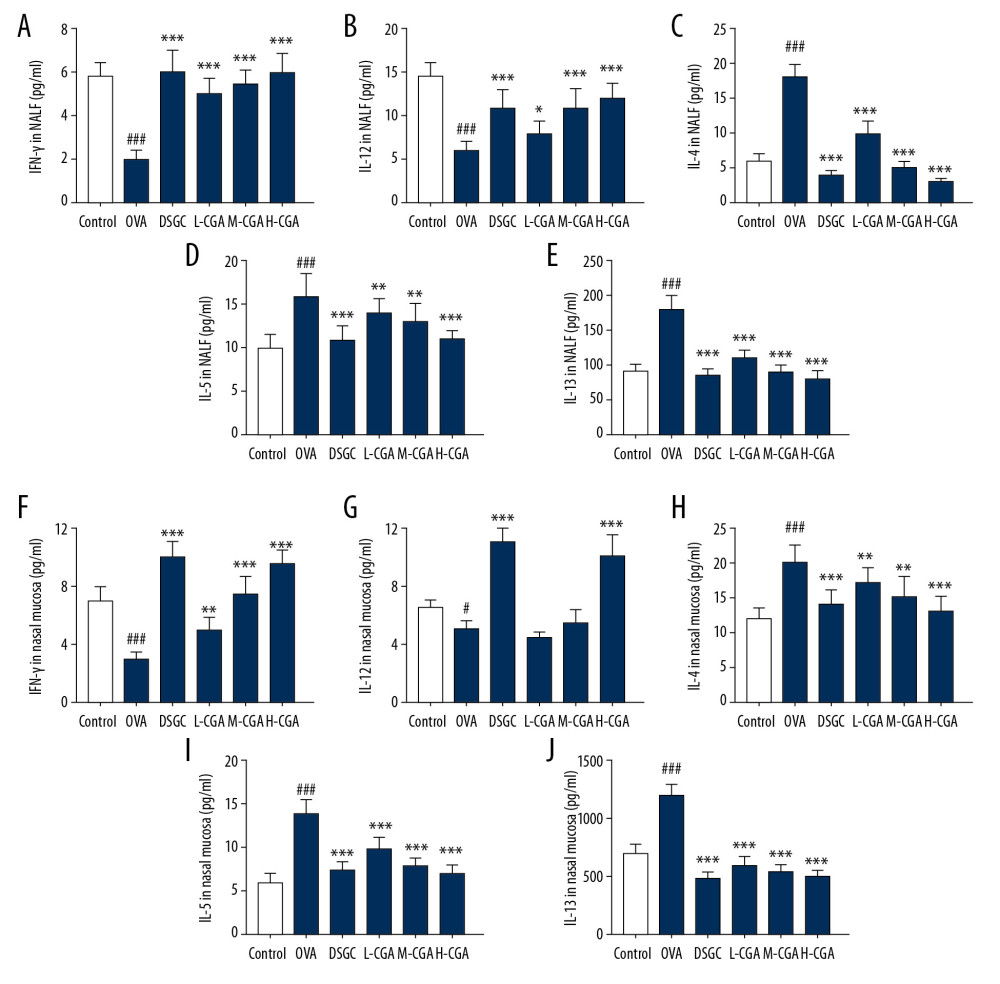 Figure 4. Chlorogenic acid (CGA) regulated the balance of Th1/Th2-associtaed cytokines in NALF and nasal tissues of the mice. Th1-related cytokine levels of IFN-γ and IL-12, and Th2-related cytokine levels IL-4, IL-5, and IL-13 were measured by enzyme-linked immunosorbent assays (ELISA) in NALF (A–E) and nasal mucosa (F–J). n=10 each group. # P<0.05, ### P<0.001 vs,. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA.
Figure 4. Chlorogenic acid (CGA) regulated the balance of Th1/Th2-associtaed cytokines in NALF and nasal tissues of the mice. Th1-related cytokine levels of IFN-γ and IL-12, and Th2-related cytokine levels IL-4, IL-5, and IL-13 were measured by enzyme-linked immunosorbent assays (ELISA) in NALF (A–E) and nasal mucosa (F–J). n=10 each group. # P<0.05, ### P<0.001 vs,. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA. 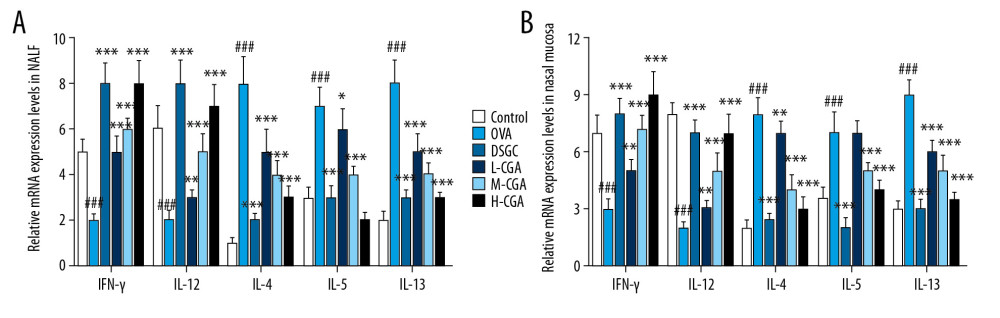 Figure 5. Chlorogenic acid (CGA) regulated mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF and nasal mucosa. Quantitative real-time polymerase chain reaction (qRT-PCR) experiments were performed to assess the mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF (A) and nasal mucosa (B). n=10 each group. ### P<0.001 vs, control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA.
Figure 5. Chlorogenic acid (CGA) regulated mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF and nasal mucosa. Quantitative real-time polymerase chain reaction (qRT-PCR) experiments were performed to assess the mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF (A) and nasal mucosa (B). n=10 each group. ### P<0.001 vs, control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA. 
References
1. Malmhall C, Bossios A, Pullerits T, Lotvall J, Effects of pollen and nasal glucocorticoid on FOXP3+, GATA-3+ and T-bet+ cells in allergic rhinitis: Allergy, 2007; 62(9); 1007-13
2. Bernstein DI, Schwartz G, Bernstein JA, Allergic rhinitis: Mechanisms and treatment: Immunol Allergy Clin, 2016; 36(2); 261-78
3. Greiner AN, Hellings PW, Rotiroti G, Scadding GK, Allergic rhinitis: Lancet (London, England), 2011; 378(9809); 2112-22
4. Kakli HA, Riley TD, Allergic rhinitis: Prim Care, 2016; 43(3); 465-75
5. Meltzer EO, Allergic rhinitis: Burden of illness, quality of life, comorbidities, and control: Immunol Allergy Clin, 2016; 36(2); 235-48
6. Maurer M, Zuberbier T, Undertreatment of rhinitis symptoms in Europe: Findings from a cross-sectional questionnaire survey: Allergy, 2007; 62(9); 1057-63
7. Castillo J, Dimov V, Investigational drugs for the treatment of allergic rhinitis: Expert Opin Investig, 2014; 23(6); 823-36
8. Zhang Y, Zhang Y, Gu W, Th1/Th2 cell’s function in immune system: Adv Exp Med Biol, 2014; 841; 45-65
9. Romagnani S, T-cell subsets (Th1 versus Th2): Ann Allergy Asthma Immunol, 2000; 85(1); 9-18 quiz 18, 21
10. Zhang Y, Zhang Y, Gu W, Sun B, TH1/TH2 cell differentiation and molecular signals: Adv Exp Med Biol, 2014; 841; 15-44
11. Bui TT, Kwon DA, Choi DW, Rosae multiflorae fructus extract and its four active components alleviate ovalbumin-induced allergic inflammatory responses via regulation of Th1/Th2 imbalance in BALB/c rhinitis mice: Phytomedicine, 2019; 55; 238-48
12. Naveed M, Hejazi V, Abbas M, Chlorogenic acid (CGA): A pharmacological review and call for further research: Biomed Pharmacother, 2018; 97; 67-74
13. Kwon SH, Lee HK, Kim JA, Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice: Eur J Pharmacol, 2010; 649(1–3); 210-17
14. Shin JY, Sohn J, Park KH, Chlorogenic acid decreases retinal vascular hyperpermeability in diabetic rat model: J Korean Med Sci, 2013; 28(4); 608-13
15. Ding Y, Cao Z, Cao L, Antiviral activity of chlorogenic acid against influenza A (H1N1/H3N2) virus and its inhibition of neuraminidase: Sci Rep, 2017; 7; 45723
16. Jaeschke H, Acetaminophen hepatotoxicity and sterile inflammation: The mechanism of protection of Chlorogenic acid: Chem Biol Interact, 2016; 243; 148-49
17. Ong KW, Hsu A, Tan BK, Anti-diabetic and anti-lipidemic effects of chlorogenic acid are mediated by ampk activation: Biochem Pharmacol, 2013; 85(9); 1341-51
18. Shin HS, Satsu H, Bae MJ, Anti-inflammatory effect of chlorogenic acid on the IL-8 production in Caco-2 cells and the dextran sulphate sodium-induced colitis symptoms in C57BL/6 mice: Food Chem, 2015; 168; 167-75
19. Chauhan PS, Satti NK, Sharma P, Differential effects of chlorogenic acid on various immunological parameters relevant to rheumatoid arthritis: Phytother Res, 2012; 26(8); 1156-65
20. Gong J, Liu FT, Chen SS, Polyphenolic antioxidants enhance IgE production: Immunol Invest, 2004; 33(3); 295-307
21. Kim HR, Lee DM, Lee SH, Chlorogenic acid suppresses pulmonary eosinophilia, IgE production, and Th2-type cytokine production in an ovalbumin-induced allergic asthma: Activation of STAT-6 and JNK is inhibited by chlorogenic acid: Int Immunopharmacol, 2010; 10(10); 1242-48
22. Tsang MS, Jiao D, Chan BC, Anti-inflammatory activities of pentaherbs formula, berberine, gallic acid and chlorogenic acid in atopic dermatitis-like skin inflammation: Molecules (Basel, Switzerland), 2016; 21(4); 519
23. Guenin S, Mauriat M, Pelloux J, Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references: J Exp Bot, 2009; 60(2); 487-93
24. Galli SJ, Tsai M, Piliponsky AM, The development of allergic inflammation: Nature, 2008; 454(7203); 445-54
25. Wallace DV, Dykewicz MS, Bernstein DI, The diagnosis and management of rhinitis: an updated practice parameter: J Allergy Clin Immunol, 2008; 122(2 Suppl); S1-84
26. Galli SJ, The mast cell-IgE paradox: From homeostasis to anaphylaxis: Am J Pathol, 2016; 186(2); 212-24
27. Pal I, Sinha Babu A, Halder I, Kumar S, Nasal smear eosinophils and allergic rhinitis: Ear Nose Throat J, 2017; 96(10–11); E17-22
28. Skoner DP, Allergic rhinitis: Definition, epidemiology, pathophysiology, detection, and diagnosis: J Allergy Clin Immunol, 2001; 108(1 Suppl); S2-8
29. Amin K, The role of mast cells in allergic inflammation: Respir Med, 2012; 106(1); 9-14
30. Yang Z, Robinson MJ, Chen X, Regulation of B cell fate by chronic activity of the IgE B cell receptor: eLife, 2016; 5; e21238
31. Kim YH, Park CS, Lim DH, Antiallergic effect of anti-Siglec-F through reduction of eosinophilic inflammation in murine allergic rhinitis: Am J Rhinol Allergy, 2013; 27(3); 187-91
32. Kak G, Raza M, Tiwari BK, Interferon-gamma (IFN-gamma): Exploring its implications in infectious diseases: Biomol Concepts, 2018; 9(1); 64-79
33. Trinchieri G, Interleukin-12 and the regulation of innate resistance and adaptive immunity: Nat Rev Immunol, 2003; 3(2); 133-46
Figures
Figure 1. Chlorogenic acid (CGA) relieved the symptom of allergic rhinitis (AR) induced by ovalbumin (OVA) in mice. (A) The chemical formula of CGA is shown in the figure. (B) The mice were sensitized on days 0, 7, and 14 by OVA and Al(OH)3, and then orally gavaged from day 15 to 25 (11 days) with CAG (50, 100, and 200 mg/kg), Cromolyn disodium salt hydrate (DSGC, 100 mg/kg), or isopycnic distilled water once a day. Next, the mice were challenged with saline containing 10% OVA via nose drops 30 minutes (min) after gavage. Before sacrifice on day 26, (C, D), nasal-rubbing and -sneezing frequencies of the mice in different groups were counted on day 25. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA.Figure 2. Chlorogenic acid (CGA) reduced the infiltration of differential inflammatory cells in nasal lavage fluid (NALF) and prevented epithelial cells disruption. NALF was collected immediately after sacrifice, and the cells were isolated by cytospin. (A) Cells from different groups were stained by Diff-Quick. Red arrows indicate eosinophils. Black arrows indicate epithelial cells lost from inflammatory tissues. (B) The number of total cells in NALF. (C) The number of eosinophils in NALF. (D) The number of neutrophils in NALF. (E) The number of lymphocytes in NALF. (F) The number of macrophages in NALF. Scale bar: 25μm, ×400 magnification. n=10 per group. ### P<0.001 vs. control, *** P<0.001 vs. OVA.Figure 3. Chlorogenic acid (CGA) reduced mucosa thickness and mucus secretions in the nasal mucosa of mice. (A) Histological features of the nasal mucosa were identified by hematoxylin-eosin (HE) staining, and the mucosa thickness of nasal septum was measured. Enzyme-linked immunosorbent assays (ELISA) were performed to detect the levels of anti-OVA specific IgE (B), anti-OVA IgG1 (C) and anti-OVA IgG2a (D) in the serum of the mice. Scale bar: 10μm, ×1000 magnification. n=10 each group. ### P<0.001 vs. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA.Figure 4. Chlorogenic acid (CGA) regulated the balance of Th1/Th2-associtaed cytokines in NALF and nasal tissues of the mice. Th1-related cytokine levels of IFN-γ and IL-12, and Th2-related cytokine levels IL-4, IL-5, and IL-13 were measured by enzyme-linked immunosorbent assays (ELISA) in NALF (A–E) and nasal mucosa (F–J). n=10 each group. # P<0.05, ### P<0.001 vs,. control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA.Figure 5. Chlorogenic acid (CGA) regulated mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF and nasal mucosa. Quantitative real-time polymerase chain reaction (qRT-PCR) experiments were performed to assess the mRNA expressions of IFN-γ, IL-12, IL-4, IL-5, and IL-13 in NALF (A) and nasal mucosa (B). n=10 each group. ### P<0.001 vs, control, * P<0.05, ** P<0.01, *** P<0.001 vs. OVA. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387