24 August 2020: Database Analysis
Expression Profile and Prognostic Values of Mini-Chromosome Maintenance Families (MCMs) in Breast Cancer
Lin Cheng1ABCDEF, Zhangmin Tan2BCDEF, Zenan Huang1BCDF, Yuhang Pan3CDF, Wenhui Zhang4CDF, Jiani Wang1ADEG*DOI: 10.12659/MSM.923673
Med Sci Monit 2020; 26:e923673






Abstract
BACKGROUND: Mini-chromosome maintenance families (MCMs) were considered the key factors for DNA replication initiation. Emerging evidences indicate that MCM2-7 (MCMs) are highly expressed in tissues from various malignant tumors. However, little is known about the clinical values of MCMs in breast cancer.
MATERIAL AND METHODS: In our study, a comprehensive bioinformatics analysis was performed to investigate expression patterns, potential functions, and prognostic values of MCMs in breast cancer, through ONCOMINE, bc-GenExMiner v4.1, Kaplan-Meier Plotter, cBioPortal and GeneMANIA databases.
RESULTS: We found that mRNA levels of MCMs were significantly elevated in breast cancer, especially in fast-growing and spreading tumor subtypes. These over-expressed MCMs predicted worse prognosis for breast cancer patients with shorter relapse-free survival (RFS) and overall survival. Among these six factors, high expression of MCM2/4/5/7 significantly reduced the RFS for patients with Luminal-A or B breast cancer and elevated MCM6/7 indicated shorter RFS for patients with basal-like or HER2-positive breast cancer. We also found that genomic alteration of MCMs was frequently found in breast cancer and the most common alteration was mRNA upregulation and amplification. Furthermore, MCMs were highly correlated with CDC45, CDC7, TIMELESS, ORC6, MCM10, ORC5, ORC4 and ORC3, mainly functioning to control the DNA replication initiation and genome stability.
CONCLUSIONS: These results suggest that MCMs are attractive prognostic biomarkers for breast cancer. Our study also provides useful clinical information about the potential of MCMs as therapeutic targets.
Keywords: Breast Neoplasms, Databases, Genetic, Ki-67 Antigen, Minichromosome Maintenance Proteins, Biomarkers, Tumor, Computational Biology, Gene Regulatory Networks, RNA, Messenger
Background
Breast cancer is the most common cancer in women and every year, nearly 1.7 million new cases are diagnosed worldwide, or 30% of all newly diagnosed cancers in women [1,2]. Although the pathogenesis of breast cancer has been extensively studied, patients who are at high risk of recurrence still suffer from low survival. Therefore, specific and sensitive markers for prognosis of breast cancer are very crucial to identify cancer patients with high risk of metastasis or recurrence, and for appropriate selection and administration of therapies for individual patients. However, conventional predictors such as tumor diameter, lymph node metastasis, histological grade, and hormone receptor and HER2 status as well as Ki-67 proliferation index seem to be insufficient for assessing risk of metastasis and prognosis in patients with breast cancer [3]. Hence, novel predictive and prognostic biomarkers are needed to identify patients who have worse prognosis and need to be treated more aggressively.
Proliferative capacity is one of the most essential features of malignant tumor [4], and proliferation of tumor cells can impact aggressiveness, clinical behavior, and prognosis of breast cancer [5]. Thus, accurate measurement of tumor proliferative capacity may not only provide useful information about patient prognosis but also help to select a more appropriate therapeutic regimen [6]. Many genes associated with DNA or cell replication are highly expressed in breast cancer tissues compared with that in normal tissues, which makes them useful biomarkers for prognosis or diagnosis of breast cancer.
The mini-chromosome maintenance family (MCM) has been considered as the key factor for DNA replication initiation [7]. The MCM family mainly consists of six highly conserved proteins, MCM2-7. Despite sharing the same MCM name, other genes including
In contrast with traditional biological assays, microarray technology allows simultaneous measurement of tens of thousands of DNA fragments for copy number variation analysis or of mRNA transcripts for gene expression [14]. As a result, microarray analysis has become one of the most important components of biomedical and biological research. Recently, the prognostic significance of several biomarkers such as PAKs, COL10A1 and BIRC5 in breast cancer has been evaluated using an online database [15–17]. In the current study, we systemically analyzed mRNA transcription levels of MCMs and their prognostic values in patients with breast cancer. Such an analysis serves to deepen our understanding of the complexity and heterogeneity of the molecular biology of breast cancer and demonstrates its clinical significance in diagnosis, prognosis, and even potentially in guiding decisions about therapy for breast cancer.
Material and Methods
ONCOMINE AND GEPIA DATABASE ANALYSIS:
ONCOMINE is an online cancer microarray database and integrated data-mining platform (www.oncomine.org) [18]. In this study, ONCOMINE 4.5 was used to analyze mRNA expression levels of MCMs in various types of breast cancers (Figure 1). Transcription levels of MCMs in cancer specimens were compared with that in normal tissue counterpart. Search criteria were for mRNA datasets and normal tissue vs. cancer tissue analysis only, and the cut-off of fold change, P value, and gene rank were defined as 2, 0.001 and the top 5%, respectively. In addition, we used the GEPIA database (http://gepia.cancer-pku.cn) [19] to analyze the hub genes expression in breast cancer (Figure 1).
BREAST CANCER GENE-EXPRESSION MINER V4.1:
bc-GenExMiner v4.1 is a statistical tool that can be used to mine 36 published annotated genomic databases of information on 4823 patients with breast cancer [20,21]. Three types of analyses are available: “expression”, “prognostic,” and “correlation” analyses, all of which have different subtypes. In this study, the expression module was used to assess expression of MCMs in breast cancer according to clinical parameters (Figure 1), such as nodal status, hormonal receptors, Scarff Bloom & Richardson grade status, and molecular subtypes.
KAPLAN-MEIER PLOTTER DATABASE ANALYSIS:
Kaplan-Meier (K-M) plotter, an open database (www.kmplot.com) including data on expression of 54 000 genes and survival information on 6234 patients with breast cancer [22], was used to assess the prognostic value of transcription levels of MCMs in breast carcinoma (Figure 1). To evaluate the prognostic significance of the target genes, samples from patients with breast carcinoma were divided into two groups (low vs. high expression) based on median gene expression. Then, we analyzed the overall survival (OS) and relapse-free survival (RFS) in patients with breast cancer with different intrinsic subtypes using K-M survival curves. The hazard ratio (HR) with 95% confidence intervals (CI) and log rank P value were calculated and displayed on the plot, and the number-at-risk was also shown below the main plot.
CBIOPORTAL DATABASE ANALYSIS AND TCGA DATA:
The cBioPortal database provides integrative analysis of large-scale cancer genomics and clinical profiles datasets from 212 cancer studies (www.cbioportal.org) [23,24]. TCGA (the Cancer Genome Atlas) includes both pathological and sequencing data on 30 different cancers [25]. In this study, the invasive breast carcinoma dataset (TCGA, Cell 2015) containing 818 samples was selected for further analysis of MCMs (Figure 1) [26]. Search parameters included putative copy-number alterations from GISTIC, mutations, mRNA expression z-scores (RNA Seq V2 RSEM, ±2), and protein expression z-scores (RPPA, ±2) with default settings.
GENEMANIA DATABASE ANALYSIS AND THE TARGET GENE FUNCTION PREDICTION:
GeneMANIA database (http://genemania.org) was used to make gene-association networks for MCMs to predict the most related genes in our gene set (Figure 1) [27]. The gene interaction network was constructed by predicted interaction, shared protein domains, physical interaction, co-expression, and pathway. The query genes were in stripped circles and the related genes were colored based on their involvement in specific processes, such as “G1/S transition of mitotic cell cycle”, “DNA replication”, “nuclear chromosome part”, “MCM complex”, “DNA helicase activity”, and “DNA replication initiation”.
STATISTICAL ANALYSIS:
Student’s
Results
MCMS WERE OVEREXPRESSED IN PATIENTS WITH BREAST CANCER:
Transcription levels of MCM2-7 in 20 types of human cancer compared to normal tissues were identified using ONCOMINE database. Figure 2A shows the percentile of highest-ranking analyses was depicted for each cancer tissue type and each MCM gene. Oncomine analysis revealed that MCMs were dramatically overexpressed in a wide variety of datasets in different cancer types (Figure 2A). For patients with breast cancer, the mRNA expression of MCM2 was significantly elevated in seven datasets. Overexpression of MCM2 was found in different types of breast carcinoma tissues, including invasive ductal, medullary, invasive lobular, and mixed lobular and ductal breast cancer (Fold changes were 2.503, 3.557, 2.120, and 2.200, respectively) (Table 1). Similarly, upregulation of MCM4 was also reported in breast cancer samples in multiple datasets (Figure 2A), with a fold change of 3.081 in ductal breast cancer [28], 3.192 in invasive ductal breast cancer, 2.048 in invasive lobular breast cancer, and 3.126 in medullary breast cancer [29]. Both MCM3 and MCM7 were only upregulated in medullary breast cancer in a dataset from the study by Curtis with fold change equaling 2.101 and 2.196, respectively [29]. Consistent with the findings above, GEPIA analysis also revealed that MCMs were more highly expressed in breast cancer tissues than in normal tissues (Figure 2B).
As shown in Table 1, the transcription level of MCM6 was higher in ductal (fold change=3.149) and medullary breast cancer (fold change=2.551) than that in normal breast tissues [29,30]. Another MCM factor with increased mRNA expression in patients with breast cancer was MCM5. The fold change was 2.622 in medullary breast cancer and 2.082 in intraductal cribriform breast adenocarcinoma compared with normal breast tissues. Of note, MCM4 and MCM5 were observed dramatically downregulated in tumor stroma relatively to control type-matched tissue, with the fold change as low as −10.866 and −4.433, respectively [31].
TRANSCRIPTION LEVELS OF MCMS WERE HIGHLY CORRELATED WITH CLINICOPATHOLOGICAL FEATURES IN PATIENTS WITH BREAST CANCER:
Based on different clinicopathological parameters, mRNA expression levels of MCMs between groups of patients were compared with bc-GenExMiner v4.1. As shown in Table 2, for age criterion, patients ≤51 yrs had higher levels of expression of MCM2/3/6/7 than those >51 yrs. As for nodal status, there was no significant difference between the two groups, except for downregulation of MCM3 in patients with negative nodes. Progesterone receptor (PR) and estrogen receptor (ER) status were observed to be negatively related with MCM2/3/4/5/6/7 expression. Conversely, mRNA levels of MCM2/3/4/5/6/7 were significantly increased in patients with triple negative breast cancer (TNBC). Furthermore, MCM2 and MCM4 were also overexpressed in patients with HER2-positive breast cancer.
As shown in Figure 3, MCM2/3/4/5/6/7 mRNA expression levels in basal-like breast cancer were the highest among the four molecular subtypes. The lowest mRNA levels of MCM2/3/4/5/6/7 were found in patients with Luminal-A breast cancer. However, no significant difference in MCM transcript levels was achieved between HER2-positive and Luminal-B subtypes of breast cancer, except for MCM2, which was upregulated in Luminal-B breast cancer. With regard to Scarff Bloom & Richardson grade status (SBR), higher mRNA levels of MCM2/3/4/5/6/7 were associated with more advanced SBR grade and all groups in the comparison (SBR3 >SBR2 >SBR1) met the cutoff, P<0.05 (Figure 4).
INCREASED MRNA EXPRESSION OF MCMS PREDICTED POORER OS AND RFS IN PATIENTS WITH BREAST CANCER:
We then assessed prognostic values of MCMs for patients with breast cancer. Kaplan-Meier plotter survival analyses and log-rank test analyses revealed that high levels of expression of MCM2/3/4/5/6/7 were significantly associated with poor OS, with HRs of 1.76, 1.29, 1.71, 1.27, 1.52, and 1.63, respectively (Figure 5). Also, RFS analysis revealed a strong correlation with all six MCM factors. As depicted in Figure 6, overexpression of MCM2/3/4/5/6/7 resulted in shorter RFS for patients with all breast cancer subtypes.
PROGNOSTIC VALUE OF MCMS IN DIFFERENT MOLECULAR SUBTYPES OF BREAST CANCER:
We further investigated the relationship between MCMs and RFS in different molecular subtypes of breast cancer. In Luminal-A subtype, high expression of MCM2/4/5/7 significantly reduced RFS in patients with breast cancer (Figure 7). A similar expression pattern was shown in patients with Luminal-B subtype breast cancer, and overexpression of MCM2/4/5/7 was also correlated with decreased RFS and worse prognosis (Figure 8). For patients with HER2-positive breast cancer, elevated MCM6/7 expression predicted shorter RFS (Figure 9). However, in the subtype with basal-like breast cancer, only patients with high expression of MCM6 presented with shorter RFS (Figure 10). Interestingly, in this study, we found that high levels of expression of MCM3 were associated with longer RFS in patients with HER2-positive breast cancer (HR=0.56 [0.38–0.83]), and high mRNA levels of MCM3/4 showed favorable RFS in patients with basal-like breast cancer (HR=0.7 [0.55–0.91] and 0.75 [0.57–0.99], respectively) (Figures 9, 10).
GENOMIC ALTERATIONS IN MCMS IN BREAST CANCER AND POTENTIAL GENE-GENE INTERACTION NETWORK FOR MCMS:
We also investigated genomic alterations in MCMs in breast cancer by examining mutational data publicly available in the cBioPortal database. The frequency of alteration in each kind of the selected genes is shown in Figure 11A. Overall, alterations in MCMs occurred in 279 of 817 sequenced invasive breast carcinoma samples (TCGA, Cell 2015), with an incidence as high as 34%. More than 45% of them (127/279) had two or more alterations. This analysis also showed that MCM4 has the highest incidence of alteration among the six MCMs (24%). The most common alteration in MCMs was mRNA upregulation and amplification.
To further explore the gene-gene interaction network and potential regulation of MCMs in breast carcinoma, we then performed data mining and constructed an interaction network using GeneMANIA online software. MCMs were found to be correlated with other genes in terms of the following additional traits: predicted interaction (36.10%), physical interaction (35.98%), shared protein domains (22.90%), co-expression (4.70%), and pathway (0.32%) (Figure 11B–11G). The network consists of 26 genes, including six identified genes and 20 additional genes that were pulled by GeneMANIA. Overall, the analysis showed that these six MCM factors interacted closely with each other. As illustrated in Figure 11C, 11E, and 11F, physical interactions, co-expression, and certain pathway are apparent between MCMs and known genes, such as CDC45, CDC7, TIMELESS, ORC6, MCM10, ORC5, ORC4, and ORC3. Among the targeted genes, the protein products of MCMs have common structural domains with MCMDC2, MCM9 and MCM8 (Figure 11D). In addition, the network in Figure 11B also predicted other functional relationships between MCMs and CDC45, CDC7, TIMELESS, ORC5, GINS4, and ORC4.
Discussion
Overexpression of and mutations in proliferation-related genes have been proved to be strongly correlated with tumor formation. Identification of novel biomarkers for cell proliferation is urgent because of their role in diagnosis, prognosis, and treatment of breast cancer, particularly for specific subtypes. MCMs were first identified in genetic screenings for
As a member of the MCM family, the role of MCM2 during cell cycle and DNA proliferation has been firmly established. In the absence of MCM2, proliferation of eukaryotic cells cannot be completed [36]. An increasing number of studies have demonstrated that MCM2 is a promising cell proliferation biomarker for a variety of cancers, such as breast, stomach, and thyroid cancers [6,7]. Furthermore, in Yousef’s study, MCM2 appeared to be an attractive alternative to Ki-67, which could help to measure cell proliferation in breast cancer and sort out Luminal-A or B subtypes of breast cancer, which have clinical features and treatment that likely are divergent [6]. Except for Ki-67, the change in cells from proliferation to differentiation or quiescent state is also accompanied by rapid disappearance of MCM3 [37]. Data also show that the decline in proliferative activity parallels decreased expression of Ki-67, whereas MCM3 starts to decrease at a later stage [37]. In contrast to Ki-67, MCM3 is significantly increased in severe dysplasia samples compared with mild dysplasia samples [38]. Therefore, for patients with oral dysplastic lesions, MCM3 may be a more sensitive proliferative biomarker than Ki-67. Similarly, Rezazadeh’s study also shows that MCM3 is a better cytologic biomarker than Ki-67 for patients with squamous cell carcinoma as MCM3 levels are significantly higher than Ki-67 in the group with cancer [39]. MCM2 and MCM3 are very important members of the MCM family. Translocation of the MCM complex from cytoplasm to nucleus and nuclear targeting of other family members are dependent upon MCM2 and MCM3 proteins.
Genetic instability plays a pivotal role in human cancer cell development [40]. Recognition of it will offer new insights into tumor pathogenesis. Mutations in MCM4 are associated with genome instability and DNA replication and repair will be perturbed in the presence of mutant MCM4 [40]. A high percentage of MCM4 expression (>70%), which is significantly higher than Ki-67, has been demonstrated to be significantly correlated with shorter survival and lymph node metastasis in esophageal adenocarcinoma [41]. MCM5, another MCM complex component, has also been demonstrated to be linked with increased proliferative rate in cells in breast cancer, gastric adenocarcinoma, and bladder cancer [42–44]. In bladder cancer cell lines, cell proliferation and invasion can be increased by estrogen receptor beta with upregulation of MCM5 expression [44]. The core of the MCM complex consists of MCM6 and MCM7 subunits. Previous studies have demonstrated that MCM6 is overexpressed in various types of cancers and plays a key role in cell proliferation [45]. The high level of expression of MCM6 in serum is a promising independent diagnostic marker for hepatocellular carcinoma, especially in aflphafetoprotein-negative patients [46]. Also, MCM6 is a potential marker for predicting shorter survival in patients with glioma [47]. However, little is known about its prognostic significance in breast cancer. Similarly, as a proliferative factor, MCM7 is correlated with tumorigenesis in a variety of malignant tumors and associated with poor prognosis [48,49]. MCM7 has been reported to be a crucial target of some oncogenic and tumor suppressor signaling pathways, which implies that multiple layers of regulation may be involved in MCM7’s biological function and its oncogenic properties [50]. One of the mechanisms is that MCM7-mediated DNA replication is potentiated by epidermal growth factor receptor through Lyn phosphorylation in human breast cancers [48]. Although these findings suggest that high levels of MCM expression tend to indicate increased biological malignant aggressiveness, other studies in various cancers, including ER-positive breast cancer, have indicated the potential role of MCM overexpression in predicting better prognosis [51]. Therefore, the prognostic significance of MCMs in breast carcinoma remains to be elucidated.
Our study showed that these six DNA replication licensing factors, MCM2-7, had distinct mRNA expression signatures in breast carcinoma tissues compared with normal tissues. MCM2-7 were significantly overexpressed in invasive lobular, medullary, invasive ductal, and mixed ductal and lobular breast cancer. The observation is noteworthy that MCM4/5 mRNA levels were dramatically downregulated in invasive breast carcinoma stromal tissue. Our analysis also indicated that mRNA levels of MCMs were significantly upregulated in younger patients with higher SBR grade status and lower expression of ER and PR. Meanwhile, the highest levels of MCM expression were found in patients with basal-like breast cancer or TNBC. These results predicted that upregulated MCMs were associated with fast-growing and spreading tumors.
In general, survival analyses revealed that higher expression of MCM2/3/4/5/6/7 was highly correlated with poorer RFS and OS in patients with all subtypes of breast cancer. Among them, higher mRNA levels of MCM2 and MCM5 mainly indicated shorter RFS for patients with Luminal-A or B breast cancer. Overexpression of MCM3 was correlated with favorable outcome for patients with basal-like or HER2-positive breast cancer. MCM4 in tumor tissue predicted shorter RFS in patients with Luminal-A or B breast cancer but longer RFS in patients with TNBC. In addition, MCM6 status was negatively correlated with RFS in patients with basal-like or HER2-positive breast cancer and MCM7 overexpression dramatically decreased the RFS probability of patients with HER2-positive, Luminal-A or Luminal-B breast cancer. Our study also revealed that MCM genomic alteration was frequently found in invasive breast carcinoma, which accounted for more than 34% of all samples. The most common alteration in MCMs was mRNA upregulation and amplification. Meanwhile, we tried to investigate the mechanisms of MCMs in regulating DNA replication origin. Results from GeneMANIA showed that MCMs were highly correlated with CDC45, CDC7, TIMELESS, ORC6, MCM10, ORC5, ORC4, and ORC3, mainly functioning to control DNA replication initiation and genome stability. Based on what we found above, MCM2-7 may be promising markers for diagnosis and prognosis of breast cancer.
As a replication protein, Ki-67 labeling index has been routinely evaluated in breast cancer. However, Ki-67 is not present in the early G1 and G0 phase of quiescent tumor cells and the level of Ki-67 may be influenced by nutrient deprivation and other external factors [43]. Besides, rather than being directly correlated with cell division, Ki-67 is considered as an “efficiency factor” in ribosome biosynthesis [52]. Therefore, the diagnostic and prognostic value of Ki-67 is limited. MCM factors, especially MCM2, MCM3 and MCM4, are more frequently expressed than standard proliferation marker Ki-67 in tumor tissues and stable throughout the cell cycle, hence, MCMs may serve as more sensitive proliferative biomarkers than Ki-67 for cytological evaluation of breast cancer [8,39,41]. In addition, it is suggested that elevated MCMs may help cancer cells resist chemotherapy and suppression of reserved MCM complexes can provide an effective sensitizing approach to increase the therapeutic index of chemotherapy drugs in cancer [53]. Therefore, elevated levels of MCMs may be potential therapeutic targets for breast carcinoma. In this study, the ONCOMINE and GEPIA databases were used to analyze expression of MCMs in breast cancer and a Kaplan-Meier plotter was used to assess prognostic values of MCMs in patients with breast cancer [18,19,22]. Furthermore, OSbrca is a novel online database for breast cancer prognostic biomarker investigation based on an independent cohort [54]. Further experiments and clinical evidence are needed to elucidate the prognostic and therapeutic value of MCMs in breast cancer.
Conclusions
In our study, a comprehensive bioinformatics analysis was performed to investigate the expression patterns, potential functions, and prognostic values of MCMs in breast carcinoma. Our results showed that MCMs were overexpressed in breast carcinoma and the overexpression predicted shorter OS and RFS for patients. High expression of MCM2/4/5/7 significantly reduced RFS in patients with Luminal-A or B breast carcinoma and elevated MCM6/7 predicted shorter RFS for patients with basal-like or HER2-positive breast carcinoma. On the one hand, MCMs might act as attractive prognostic biomarkers for breast carcinoma. In addition, effective therapeutic targets are essential for treatment of breast carcinoma. Our results provide useful clinical information about the potential of MCMs as therapeutic targets.
Figures
 Figure 1. The databases used in this study.
Figure 1. The databases used in this study. 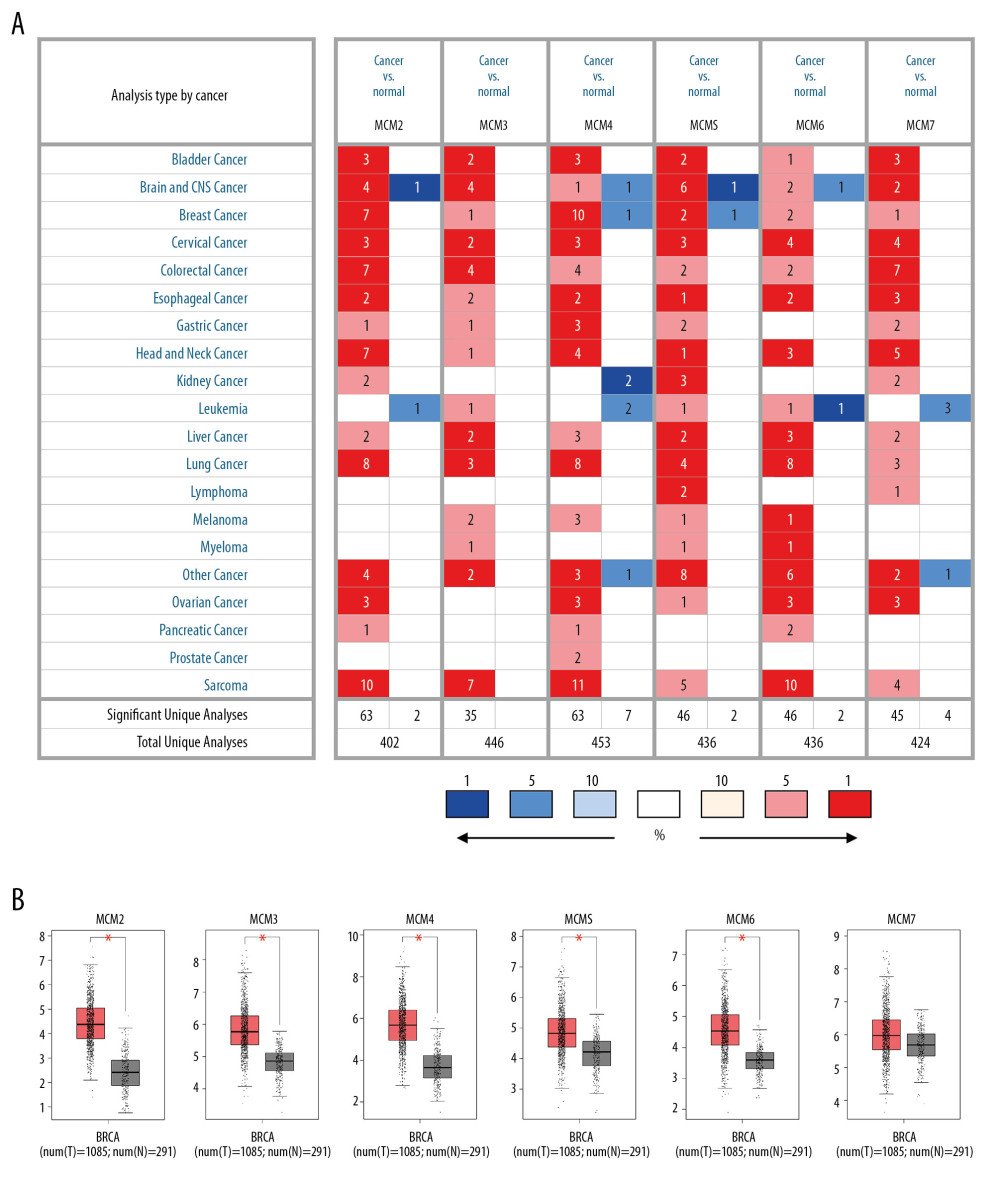 Figure 2. Expression of MCM genes across different types of cancers (A) Transcription levels of MCMs in twenty human cancers compared with normal tissue were obtained from ONCOMINE, with the gene rank ≤ top 5%, fold change ≥2 and threshold of P≤0.001. Significant mRNA underexpression or overexpression of MCMs was shown as blue or red cell. Cancer types are organized by their tissue of origin, and the degree of color was determined by the gene rank percentile of the highest-ranking analyses. (B) MCMs expression levels in breast cancer compared with normal tissues in the GEPIA database.
Figure 2. Expression of MCM genes across different types of cancers (A) Transcription levels of MCMs in twenty human cancers compared with normal tissue were obtained from ONCOMINE, with the gene rank ≤ top 5%, fold change ≥2 and threshold of P≤0.001. Significant mRNA underexpression or overexpression of MCMs was shown as blue or red cell. Cancer types are organized by their tissue of origin, and the degree of color was determined by the gene rank percentile of the highest-ranking analyses. (B) MCMs expression levels in breast cancer compared with normal tissues in the GEPIA database. 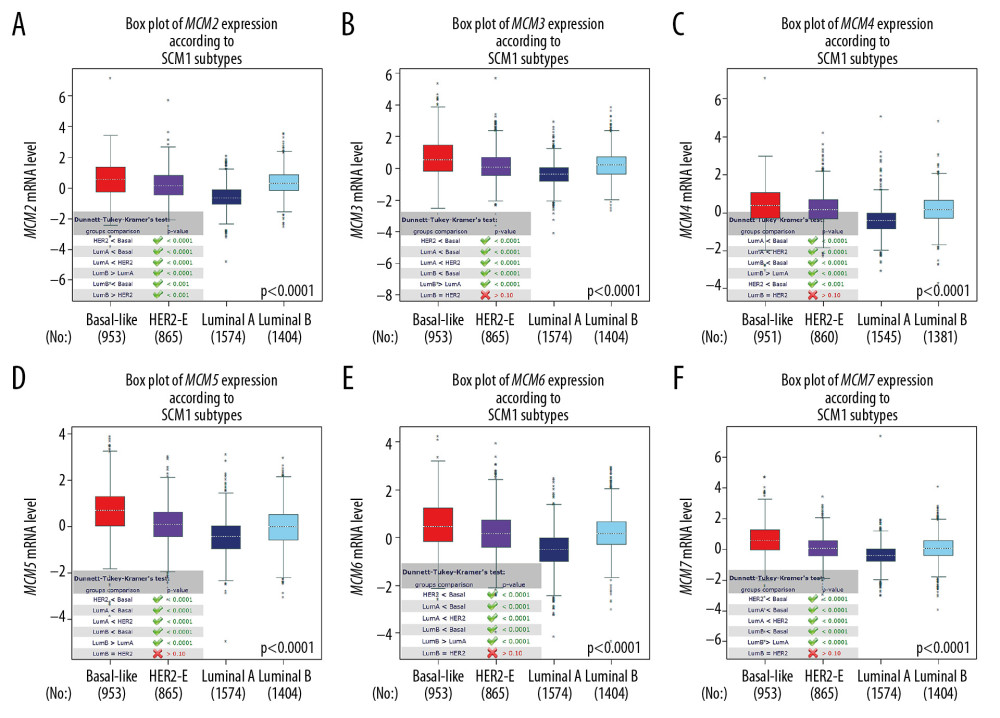 Figure 3. (A–F) mRNA expression levels of MCMs in four molecular subtypes of breast cancer. The Welch’s test was performed along with Dunnett-Tukey-Kramer’s tests for pairwise comparison when a global significant difference existed (P<0.05).
Figure 3. (A–F) mRNA expression levels of MCMs in four molecular subtypes of breast cancer. The Welch’s test was performed along with Dunnett-Tukey-Kramer’s tests for pairwise comparison when a global significant difference existed (P<0.05). 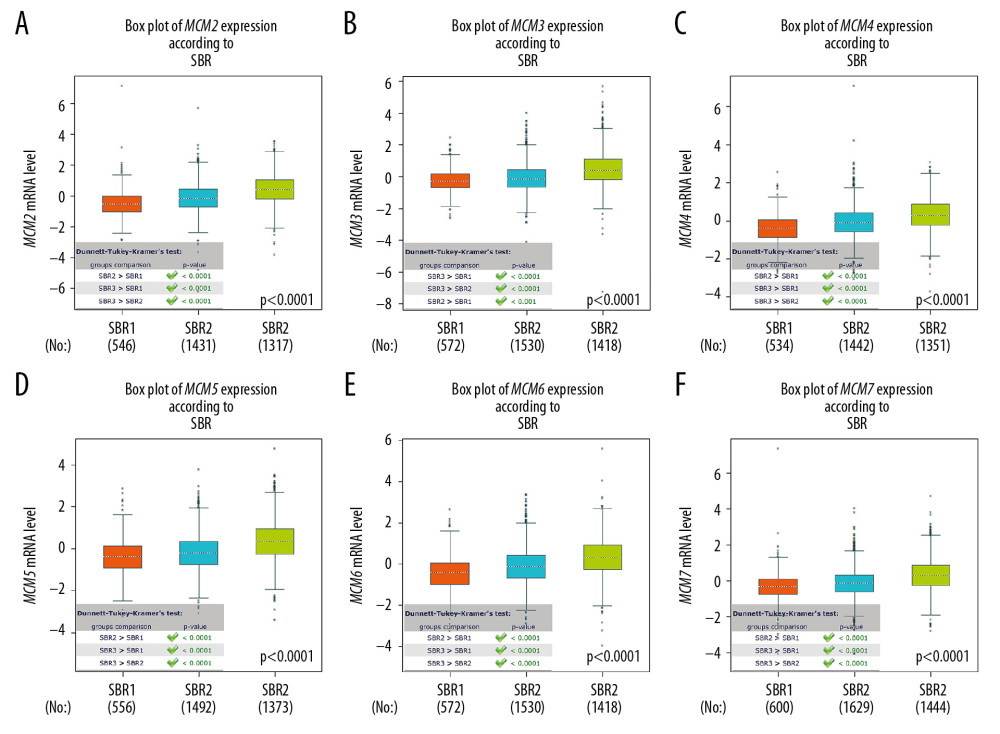 Figure 4. (A–F) mRNA expression levels of MCMs in different Scarff Bloom & Richardson grade status (SBR).
Figure 4. (A–F) mRNA expression levels of MCMs in different Scarff Bloom & Richardson grade status (SBR). 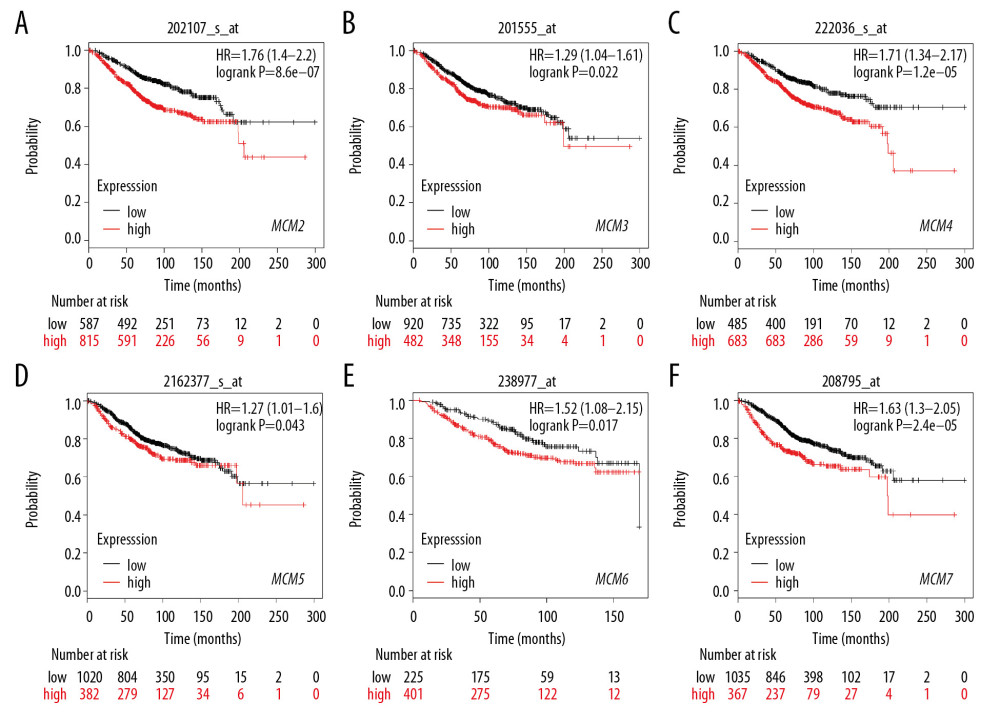 Figure 5. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (OS in Kaplan-Meier plotter).
Figure 5. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (OS in Kaplan-Meier plotter). 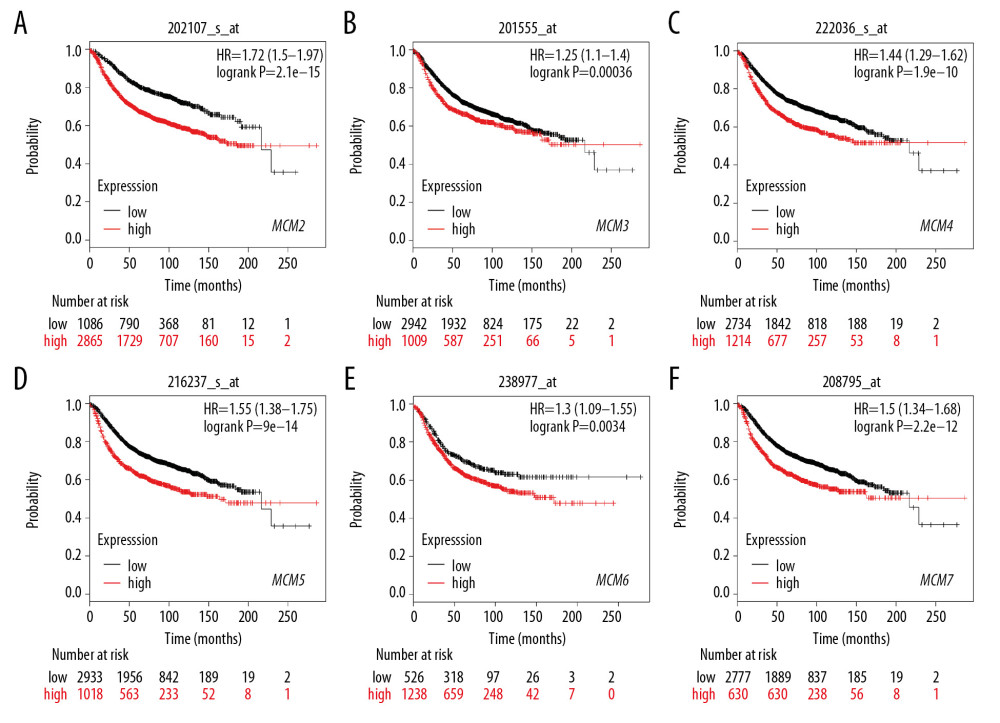 Figure 6. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (RFS in Kaplan-Meier plotter).
Figure 6. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (RFS in Kaplan-Meier plotter). 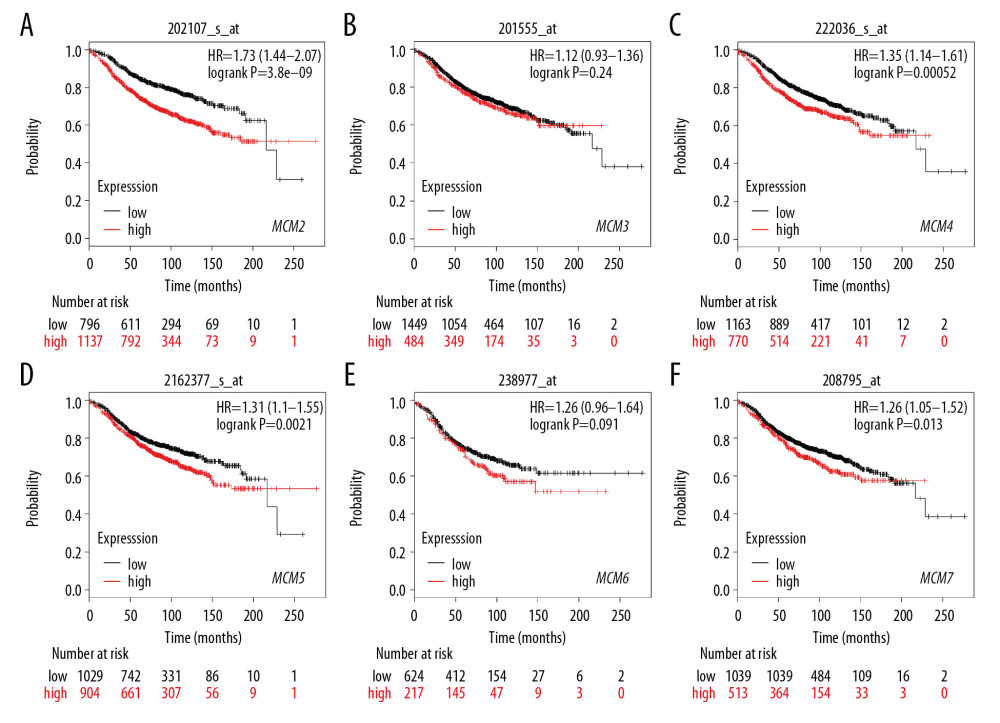 Figure 7. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-A breast cancer (RFS in Kaplan-Meier plotter).
Figure 7. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-A breast cancer (RFS in Kaplan-Meier plotter). 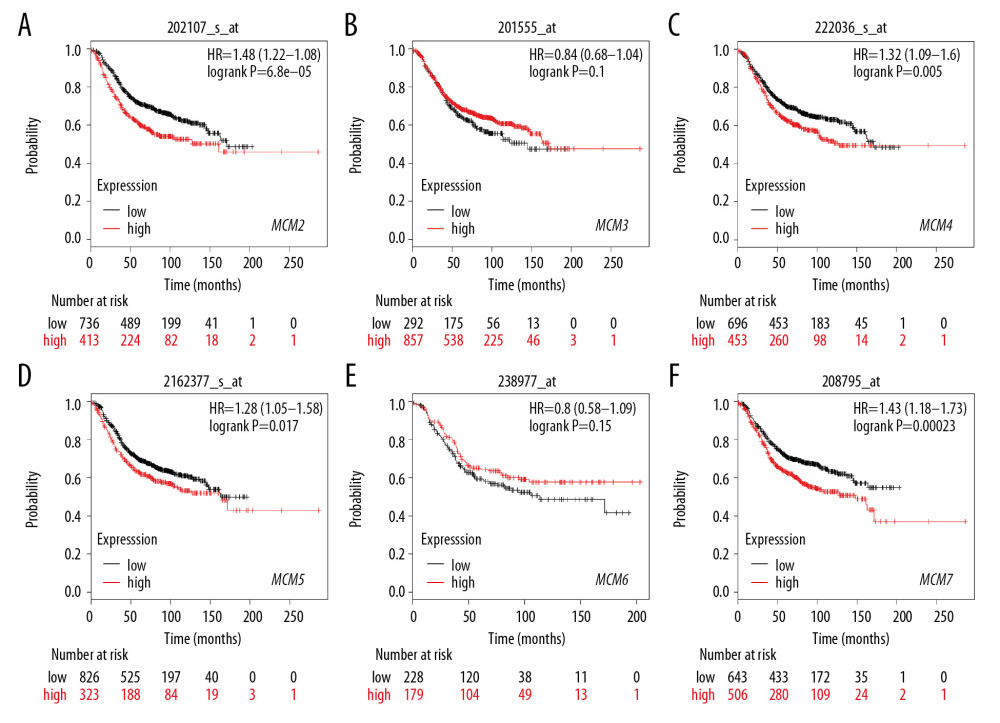 Figure 8. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-B breast cancer (RFS in Kaplan-Meier plotter).
Figure 8. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-B breast cancer (RFS in Kaplan-Meier plotter). 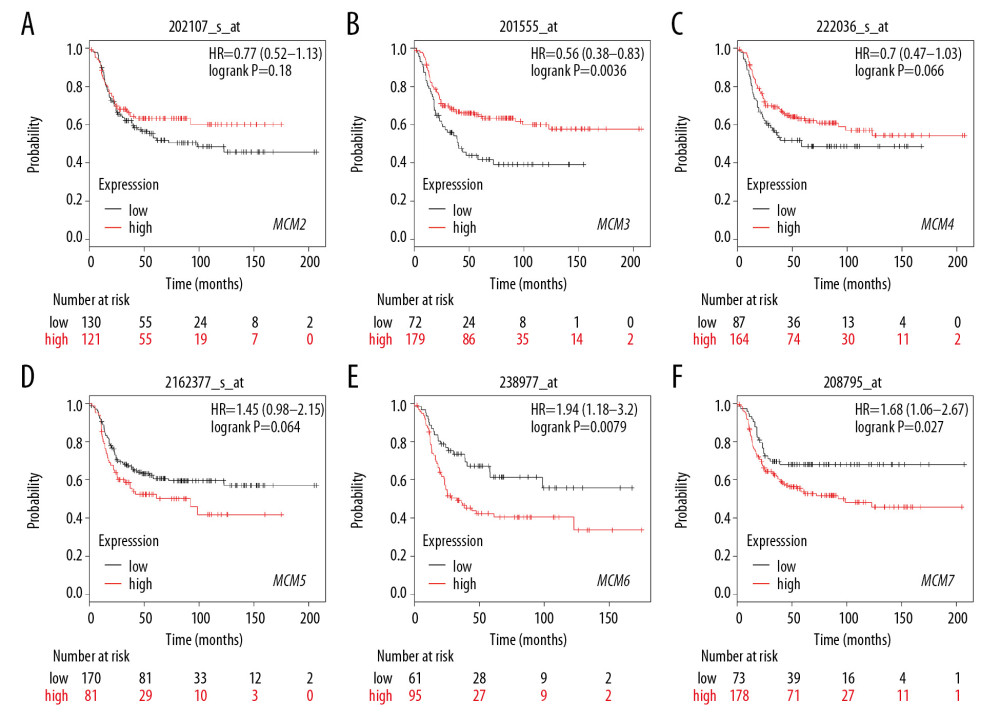 Figure 9. (A–F) Prognostic values of mRNA levels of MCMs in patients with HER2-positive breast cancer (RFS in Kaplan-Meier plotter).
Figure 9. (A–F) Prognostic values of mRNA levels of MCMs in patients with HER2-positive breast cancer (RFS in Kaplan-Meier plotter). 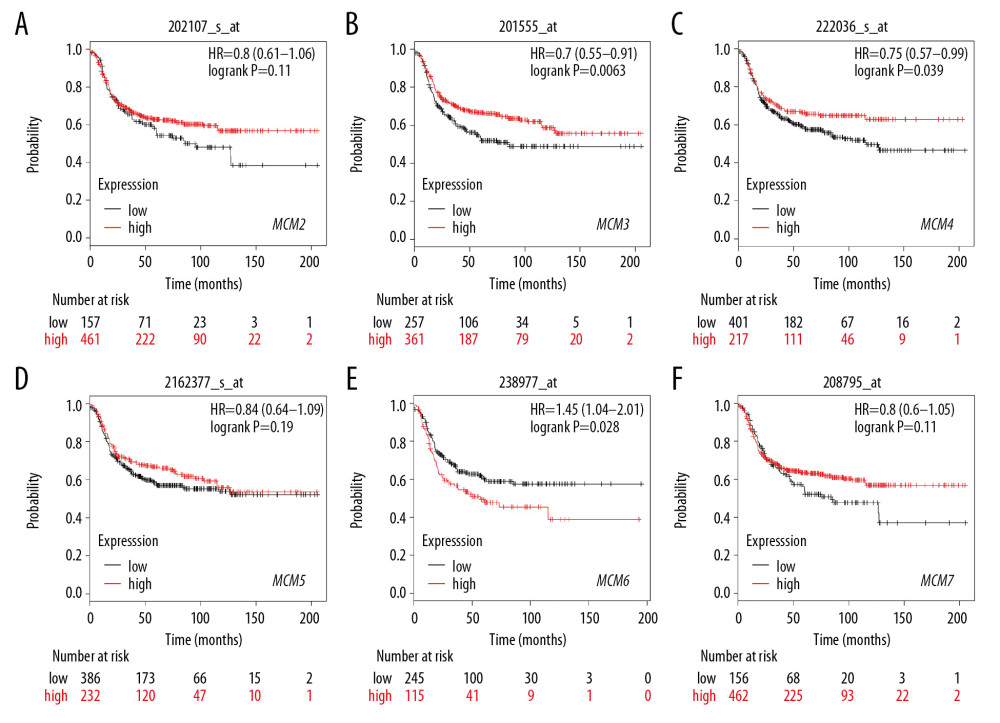 Figure 10. (A–F) Prognostic values of mRNA levels of MCMs in patients with basal-like breast cancer (RFS in Kaplan-Meier plotter).
Figure 10. (A–F) Prognostic values of mRNA levels of MCMs in patients with basal-like breast cancer (RFS in Kaplan-Meier plotter). 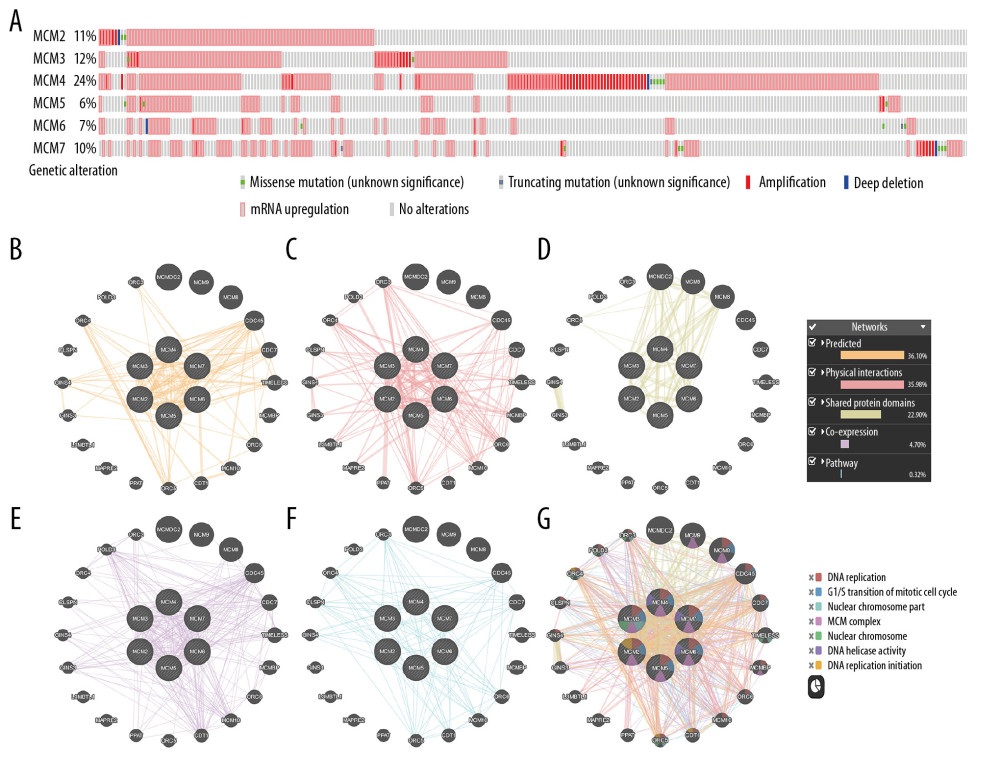 Figure 11. Genomic alterations of MCMs in breast cancer (cBioportal) and potential gene-gene interaction network for MCMs with paths colored according to their functions (GeneMANIA). (A) Distribution and proportion of MCMs alterations in breast cancer samples were displayed above. (A) Samples without alterations on the right side were cropped from the figure. G: The integrated interaction network of MCMs with other related genes was constructed by B: predicted interaction, C: physical interaction, D: shared protein domains, E: co-expression, and F: pathway.
Figure 11. Genomic alterations of MCMs in breast cancer (cBioportal) and potential gene-gene interaction network for MCMs with paths colored according to their functions (GeneMANIA). (A) Distribution and proportion of MCMs alterations in breast cancer samples were displayed above. (A) Samples without alterations on the right side were cropped from the figure. G: The integrated interaction network of MCMs with other related genes was constructed by B: predicted interaction, C: physical interaction, D: shared protein domains, E: co-expression, and F: pathway. References
1. Siegel RL, Miller KD, Jemal A, Cancer statistics, 2018: Cancer J Clin, 2018; 68(1); 7-30
2. Siegel RL, Miller KD, Jemal A, Cancer statistics, 2016: Cancer J Clin, 2016; 66(1); 7-30
3. Guler EN, Gene expression profiling in breast cancer and its effect on therapy selection in early-stage breast cancer: Eur J Breast Health, 2017; 13(4); 168-74
4. Hanahan D, Weinberg RA, Hallmarks of cancer: The next generation: Cell, 2011; 144(5); 646-74
5. Aleskandarany MA, Green AR, Benhasouna AA, Prognostic value of proliferation assay in the luminal, HER2-positive, and triple-negative biologic classes of breast cancer: Breast Cancer Res, 2012; 14(1); R3
6. Yousef EM, Furrer D, Laperriere DL, MCM2: An alternative to Ki-67 for measuring breast cancer cell proliferation: Mod Pathol, 2017; 30(5); 682-97
7. Abe S, Yamamoto K, Kurata M, Targeting MCM2 function as a novel strategy for the treatment of highly malignant breast tumors: Oncotarget, 2015; 6(33); 34892-909
8. Jurikova M, Danihel L, Polak S, Ki67, PCNA, and MCM proteins: Markers of proliferation in the diagnosis of breast cancer: Acta Histochem, 2016; 118(5); 544-52
9. Li J, Liu J, Liang Z, Simvastatin and Atorvastatin inhibit DNA replication licensing factor MCM7 and effectively suppress RB-deficient tumors growth: Cell Death Dis, 2017; 8(3); e2673
10. Wharton SB, Hibberd S, Eward KL, DNA replication licensing and cell cycle kinetics of oligodendroglial tumours: Br J Cancer, 2004; 91(2); 262-69
11. Gonzalez MA, Tachibana KE, Laskey RA, Control of DNA replication and its potential clinical exploitation: Nat Rev Cancer, 2005; 5(2); 135-41
12. Ishimi Y, Okayasu I, Kato C, Enhanced expression of MCM proteins in cancer cells derived from uterine cervix: Eur J Biochem, 2003; 270(6); 1089-101
13. Alison MR, Hunt T, Forbes SJ, Minichromosome maintenance (MCM) proteins may be pre-cancer markers: Gut, 2002; 50(3); 290-91
14. Sealfon SC, Chu TT, RNA and DNA microarrays: Methods Mol Biol, 2011; 671; 3-34
15. Dang Y, Guo Y, Ma X, Systemic analysis of the expression and prognostic significance of PAKs in breast cancer: Genomics, 2020; 112(3); 2433-44
16. Zhang M, Chen H, Wang M, Bioinformatics analysis of prognostic significance of COL10A1 in breast cancer: Biosci Rep, 2020; 40(2) BSR20193286
17. Dai JB, Zhu B, Lin WJ, Identification of prognostic significance of BIRC5 in breast cancer using integrative bioinformatics analysis: Biosci Rep, 2020; 40(2) BSR20193678
18. Rhodes DR, Yu J, Shanker K, ONCOMINE: A cancer microarray database and integrated data-mining platform: Neoplasia, 2004; 6(1); 1-6
19. Tang Z, Li C, Kang B, GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses: Nucleic Acids Res, 2017; 45(W1); W98-102
20. Jezequel P, Campone M, Gouraud W, bc-GenExMiner: An easy-to-use online platform for gene prognostic analyses in breast cancer: Breast Cancer Res Treat, 2012; 131(3); 765-75
21. Jezequel P, Frenel JS, Campion L, bc-GenExMiner 30: New mining module computes breast cancer gene expression correlation analyses: Database (Oxford), 2013; 2013 bas060
22. Gyorffy B, Lanczky A, Eklund AC, An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients: Breast Cancer Res Treat, 2010; 123(3); 725-31
23. Gao J, Aksoy BA, Dogrusoz U, Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal: Sci Signal, 2013; 6(269); pl1
24. Cerami E, Gao J, Dogrusoz U, The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data: Cancer Discov, 2012; 2(5); 401-4
25. Cancer Genome Atlas Network, Comprehensive molecular portraits of human breast tumours: Nature, 2012; 490(7418); 61-70
26. Ciriello G, Gatza ML, Beck AH, Comprehensive molecular portraits of invasive lobular breast cancer: Cell, 2015; 163(2); 506-19
27. Warde-Farley D, Donaldson SL, Comes O, The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function: Nucleic Acids Res, 2010; 38(Web Server issue); W214-20
28. Perou CM, Sorlie T, Eisen MB, Molecular portraits of human breast tumours: Nature, 2000; 406(6797); 747-52
29. Curtis C, Shah SP, Chin SF, The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups: Nature, 2012; 486(7403); 346-52
30. Richardson AL, Wang ZC, De Nicolo A, X chromosomal abnormalities in basal-like human breast cancer: Cancer Cell, 2006; 9(2); 121-32
31. Finak G, Bertos N, Pepin F, Stromal gene expression predicts clinical outcome in breast cancer: Nat Med, 2008; 14(5); 518-27
32. Simon NE, Schwacha A, The Mcm2–7 replicative helicase: A promising chemotherapeutic target: Biomed Res Int, 2014; 2014 549719
33. Liu Z, Li J, Chen J, MCM family in HCC: MCM6 indicates adverse tumor features and poor outcomes and promotes S/G2 cell cycle progression: BMC Cancer, 2018; 18(1); 200
34. Forsburg SL, Eukaryotic MCM proteins: Beyond replication initiation: Microbiol Mol Biol Rev, 2004; 68(1); 109-31
35. Stoeber K, Tlsty TD, Happerfield L, DNA replication licensing and human cell proliferation: J Cell Sci, 2001; 114(Pt 11); 2027-41
36. Laskey R, The Croonian Lecture 2001 hunting the antisocial cancer cell: MCM proteins and their exploitation: Philos Trans R Soc Lond B Biol Sci, 2005; 360(1458); 1119-32
37. Endl E, Kausch I, Baack M, The expression of Ki-67, MCM3, and p27 defines distinct subsets of proliferating, resting, and differentiated cells: J Pathol, 2001; 195(4); 457-62
38. Lameira AG, Pontes FS, Guimaraes DM, MCM3 could be a better marker than Ki-67 for evaluation of dysplastic oral lesions: An immunohistochemical study: J Oral Pathol Med, 2014; 43(6); 427-34
39. Rezazadeh F, Ebrahimi R, Andisheh-Tadbir A, Evaluation of the Ki-67 and MCM3 expression in cytologic smear of oral squamous cell carcinoma: J Dent (Shiraz), 2017; 18(3); 207-11
40. Tatsumi R, Ishimi Y, An MCM4 mutation detected in cancer cells affects MCM4/6/7 complex formation: J Biochem, 2017; 161(3); 259-68
41. Choy B, LaLonde A, Que J, MCM4 and MCM7, potential novel proliferation markers, significantly correlated with Ki-67, Bmi1, and cyclin E expression in esophageal adenocarcinoma, squamous cell carcinoma, and precancerous lesions: Hum Pathol, 2016; 57; 126-35
42. Eissa S, Matboli M, Shehata HH, MicroRNA-10b and minichromosome maintenance complex component 5 gene as prognostic biomarkers in breast cancer: Tumour Biol, 2015; 36(6); 4487-94
43. Giaginis C, Giagini A, Tsourouflis G, MCM-2 and MCM-5 expression in gastric adenocarcinoma: clinical significance and comparison with Ki-67 proliferative marker: Dig Dis Sci, 2011; 56(3); 777-85
44. Hsu I, Chuang KL, Slavin S, Suppression of ERbeta signaling via ERbeta knockout or antagonist protects against bladder cancer development: Carcinogenesis, 2014; 35(3); 651-61
45. Liu M, Hu Q, Tu M, MCM6 promotes metastasis of hepatocellular carcinoma via MEK/ERK pathway and serves as a novel serum biomarker for early recurrence: J Exp Clin Cancer Res, 2018; 37(1); 10
46. Zheng T, Chen M, Han S, Plasma minichromosome maintenance complex component 6 is a novel biomarker for hepatocellular carcinoma patients: Hepatol Res, 2014; 44(13); 1347-56
47. Cai HQ, Cheng ZJ, Zhang HP, Overexpression of MCM6 predicts poor survival in patients with glioma: Hum Pathol, 2018; 78; 182-87
48. Huang TH, Huo L, Wang YN, Epidermal growth factor receptor potentiates MCM7-mediated DNA replication through tyrosine phosphorylation of Lyn kinase in human cancers: Cancer Cell, 2013; 23(6); 796-810
49. Toyokawa G, Masuda K, Daigo Y, Minichromosome maintenance protein 7 is a potential therapeutic target in human cancer and a novel prognostic marker of non-small cell lung cancer: Mol Cancer, 2011; 10; 65
50. Han YC, Yu YP, Nelson J, Interaction of integrin-linked kinase and miniature chromosome maintenance 7-mediating integrin {alpha}7 induced cell growth suppression: Cancer Res, 2010; 70(11); 4375-84
51. Gou K, Liu J, Feng X, Expression of minichromosome maintenance proteins (MCM) and cancer prognosis: A meta-analysis: J Cancer, 2018; 9(8); 1518-26
52. MacCallum DE, Hall PA, The location of pKi67 in the outer dense fibrillary compartment of the nucleolus points to a role in ribosome biogenesis during the cell division cycle: J Pathol, 2000; 190(5); 537-44
53. Bryant VL, Elias RM, McCarthy SM, Suppression of reserve MCM complexes chemosensitizes to gemcitabine and 5-fluorouracil: Mol Cancer Res, 2015; 13(9); 1296-305
54. Yan Z, Wang Q, Sun X, OSbrca: A Web Server for breast cancer prognostic biomarker investigation with massive data from tens of cohorts: Front Oncol, 2019; 9; 1349
Figures
Figure 1. The databases used in this study.Figure 2. Expression of MCM genes across different types of cancers (A) Transcription levels of MCMs in twenty human cancers compared with normal tissue were obtained from ONCOMINE, with the gene rank ≤ top 5%, fold change ≥2 and threshold of P≤0.001. Significant mRNA underexpression or overexpression of MCMs was shown as blue or red cell. Cancer types are organized by their tissue of origin, and the degree of color was determined by the gene rank percentile of the highest-ranking analyses. (B) MCMs expression levels in breast cancer compared with normal tissues in the GEPIA database.Figure 3. (A–F) mRNA expression levels of MCMs in four molecular subtypes of breast cancer. The Welch’s test was performed along with Dunnett-Tukey-Kramer’s tests for pairwise comparison when a global significant difference existed (P<0.05).Figure 4. (A–F) mRNA expression levels of MCMs in different Scarff Bloom & Richardson grade status (SBR).Figure 5. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (OS in Kaplan-Meier plotter).Figure 6. (A–F) Prognostic values of mRNA levels of MCMs in patients under all breast cancer subtypes (RFS in Kaplan-Meier plotter).Figure 7. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-A breast cancer (RFS in Kaplan-Meier plotter).Figure 8. (A–F) Prognostic values of mRNA levels of MCMs in patients with Luminal-B breast cancer (RFS in Kaplan-Meier plotter).Figure 9. (A–F) Prognostic values of mRNA levels of MCMs in patients with HER2-positive breast cancer (RFS in Kaplan-Meier plotter).Figure 10. (A–F) Prognostic values of mRNA levels of MCMs in patients with basal-like breast cancer (RFS in Kaplan-Meier plotter).Figure 11. Genomic alterations of MCMs in breast cancer (cBioportal) and potential gene-gene interaction network for MCMs with paths colored according to their functions (GeneMANIA). (A) Distribution and proportion of MCMs alterations in breast cancer samples were displayed above. (A) Samples without alterations on the right side were cropped from the figure. G: The integrated interaction network of MCMs with other related genes was constructed by B: predicted interaction, C: physical interaction, D: shared protein domains, E: co-expression, and F: pathway. Tables
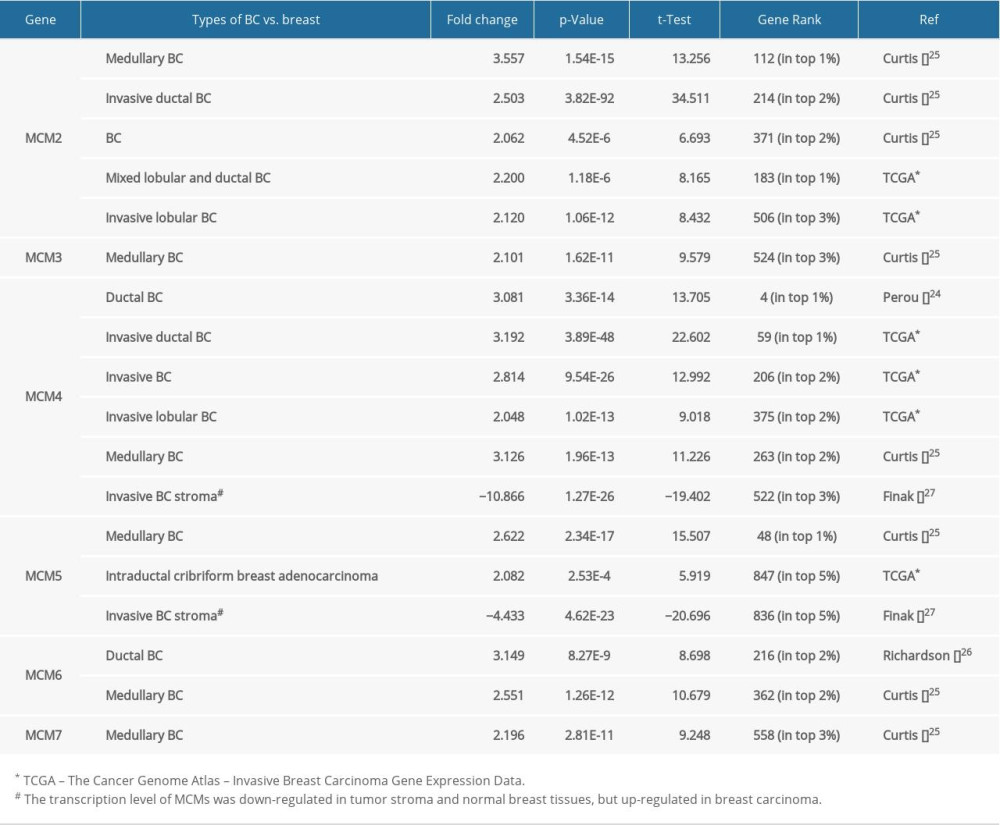 Table 1. The mRNA transcription level of MCMs in different types of breast carcinoma (BC) compared with normal breast tissues (ONCOMINE database).
Table 1. The mRNA transcription level of MCMs in different types of breast carcinoma (BC) compared with normal breast tissues (ONCOMINE database).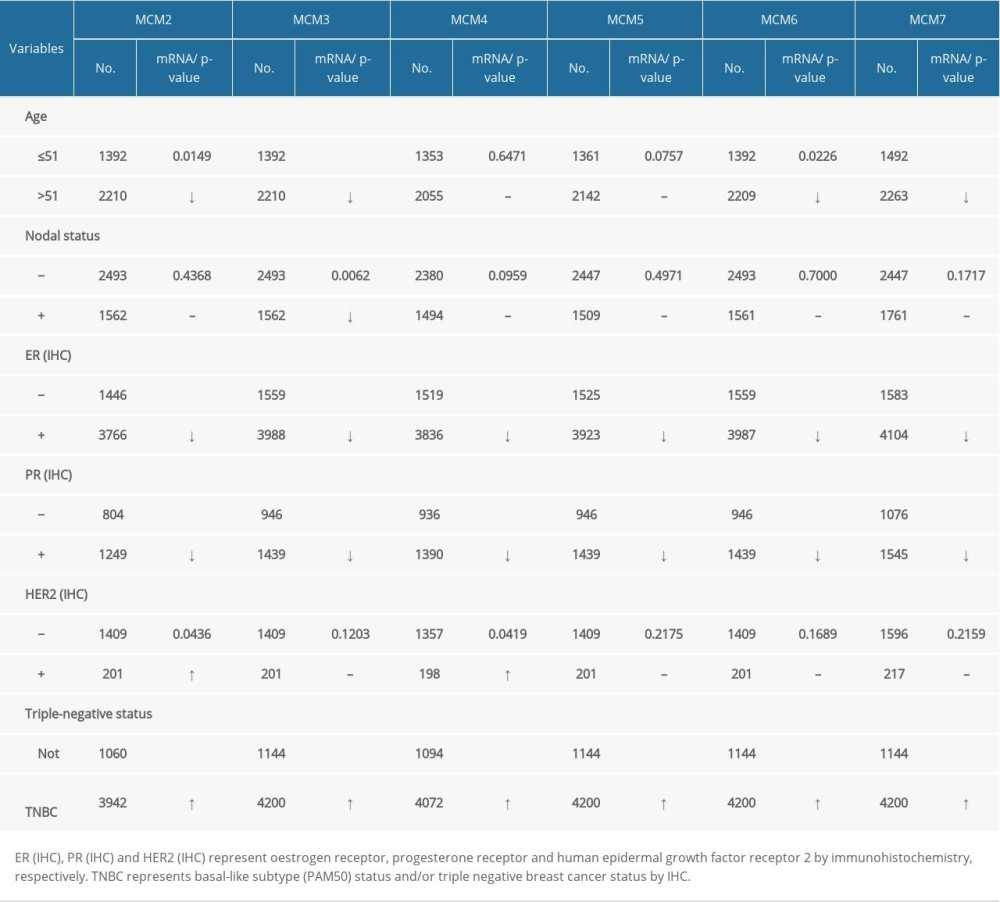 Table 2. Correlation of MCMs mRNA expression with clinicopathological features for patients with breast carcinoma.Table 1. The mRNA transcription level of MCMs in different types of breast carcinoma (BC) compared with normal breast tissues (ONCOMINE database).Table 2. Correlation of MCMs mRNA expression with clinicopathological features for patients with breast carcinoma.
Table 2. Correlation of MCMs mRNA expression with clinicopathological features for patients with breast carcinoma.Table 1. The mRNA transcription level of MCMs in different types of breast carcinoma (BC) compared with normal breast tissues (ONCOMINE database).Table 2. Correlation of MCMs mRNA expression with clinicopathological features for patients with breast carcinoma. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387