01 August 2020: Animal Study
Effects of Jian-Pi-Zhi-Dong Decoction on the Expression of 5-HT and Its Receptor in a Rat Model of Tourette Syndrome and Comorbid Anxiety
Dan Wang1BE, Hui-ling Tian2CF, Xia Cui3DG, Qian Wang3C, Fan Guo3BC, Wen Zhang3ABCDEFG*, Qi-sheng Tang1AGDOI: 10.12659/MSM.924658
Med Sci Monit 2020; 26:e924658






Abstract
BACKGROUND: Anxiety is one of the common comorbidities of Tourette syndrome (TS). The serotonin (5-HT) system is involved in both TS and anxiety. Jian-pi-zhi-dong decoction (JPZDD) is widely used. However, the mechanism remains unknown. In this study, a rat model of TS and comorbid anxiety was used to evaluate the effect of JPZDD on 5-HT and its receptor.
MATERIAL AND METHODS: 48 rats were divided into 4 groups randomly (n=12). The model was established by empty water bottle stimulation plus iminodipropionitrile injection for 3 weeks. Then the control and model groups were gavaged with saline, while the treatment groups were gavaged with fluoxetine hydrochloride (Flx) or JPZDD. Body weights were measured, and behavioral tests were evaluated with stereotypy and elevated plus maze. The morphologic characters were observed by hematoxylin and eosin staining. The content of 5-HT was detected by enzyme-linked immunosorbent assay and high-performance liquid chromatography. The expression of 5-HT2C receptor was detected by western blot and quantitative polymerase chain reaction.
RESULTS: The stereotypy score was lower and the time spent in the open arm was longer in the JPZDD group compared with the model group. After the treatment of Flx or JPZDD, the structure of neurons became gradually normal and the cells were arranged neatly. The contents of 5-HT in the treatment groups were higher compared with the model group in the striatum. The expression of 5-HT2C mRNA in the striatum of JPZDD and Flx groups decreased compared with the model group, and the JPZDD group was lower than the Flx group.
CONCLUSIONS: JPZDD alleviated both tic and anxiety symptoms and the mechanism may be via reducing the expression of 5-HT2C mRNA in the striatum, increasing the concentration of 5-HT, and enhancing the activity of the 5-HT system, which in turn exerts neuro-inhibition.
Keywords: Anxiety, Comorbidity, Neural Pathways, Receptor, Serotonin, 5-HT2C, Tourette Syndrome, Antidepressive Agents, Second-Generation, Behavior, Animal, Cerebral Cortex, Corpus Striatum, Drugs, Chinese Herbal, fluoxetine, Maze Learning, Nitriles, Serotonin, Serotonin 5-HT1 Receptor Antagonists
Background
According to the DSM-5 criteria, the presence of motor and vocal (or phonic) tics manifesting before the age of 18 years for more than 12 months in the absence of secondary causes warrants a diagnosis of Gilles de la Tourette Syndrome(TS) [1]. TS is complicated by psychiatric comorbidities (attention-deficit/hyperactivity disorder [ADHD], obsessive-compulsive disorder [OCD], anxiety/depressive disorders [AD, DD], autism spectrum disorders [ASD]) in about 90% of cases [2]. Comorbid condition refers to: 1) a potential common cause leads to 2 or more different diseases; 2) one disease causes another disease; or 3) 2 unrelated diseases occur simultaneously [3], among which anxiety is one of the common comorbidities of tics, as its incidence is just below that of ADHD and OCD [4]. The foreign (non-Chinese) literature reports that the incidence of anxiety in TS was 6.1–50.91% [5,6], with clinical manifestations in those who were fidgety, grumpy, upset, disobedient, crying, nervous, afraid, difficult to placate, complaining, losing their tempers, and groaning. Comorbid anxiety often causes more serious damage than TS itself. The degree of anxiety was correlated with the degree of tic. A higher degree of anxiety was associated with more severe tic symptoms [4]. It seriously affects the physical and mental health of children, which can be a heavy burden for families and society. Thus, the comorbidity of TS has attracted the attention of many scholars.
Currently, TS is thought to arise as the result of fundamental alternations in the functional dynamics of cortico-striato-thalamo-cortical (CSTC) pathways, which link specific regions in the frontal cortex to subcortical structures (including the basal ganglia) [7]. Imbalance of inhibitory and excitatory neurotransmitters or a combination of neurotransmitters, including dopamine (DA), serotonin (5-HT), glutamate (Glu), and γ-aminobutyric acid (GABA), located in these pathways are relevant in TS and comorbid pathogenesis. Neurobiochemical studies have found that the pathogenesis of TS is related to the hyperfunction of DA in the basal ganglia, thalamus, and cortex [8]. The 5-HT system is accompanied by the DA system, which indirectly controls the body’s movement, cognition, and emotion and participates in the occurrence of TS [9]. When 5-HT is released into the synaptic cleft, it will connect with the 5-HT receptors in the postsynaptic membrane to exert biological effects. The role of 5-HT and its receptors in the regulation of anxiety has been widely acknowledged [10]. At the same time, the 5-HT reuptake inhibitor, fluoxetine, is selected as the first-line drug to treat TS and comorbid anxiety disorders [11]. So, TS and comorbid anxiety are closely related to 5-HT metabolism.
Within the framework of traditional Chinese medicine (TCM) theory, TS originated in the spleen, concurrent to the liver, imbalance in the brain, brain spirit disturbed is the core pathogenesis, the comorbidity of anxiety is induced by uneasy of heart-shen. The therapy for treating TS and comorbid anxiety is invigorating the spleen and smoothing the liver so as to calm heart-shen [12]. Jian-pi-zhi-dong decoction (JPZDD), a Chinese herbal formulation has been clinically prescribed for the treatment of TS and comorbidities in children for about 20 years in China. It is effectively used to nourish the spleen and smooth the liver to relieve convulsions. A series of studies on the treatment of 904 cases of multiple tic symptom comorbidities using JPZDD, found that JPZDD could not only improve the symptoms of children with tics, but that it could also alleviate the symptoms of comorbidities [13]. However, the biochemical mechanism underlying the beneficial effects of JPZDD in TS and comorbid anxiety remains largely unknown. In this study, the TS and comorbid anxiety model was established by an intraperitoneal iminodipropionitrile (IDPN) injection combined with uncertain empty bottle stimulation [14], then the relationship between the behavioral changes and 5-HT content and its receptor expressions in the cortex and striatum were examined to further explore the therapeutic multiple target of JPZDD.
Material and Methods
ANIMALS:
Male Sprague Dawley rats (n=48, 50±10 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, People’s Republic of China; No SCXK 2012-0001). The animals were maintained at 21±1°C in a standard 12-hour light/dark cycle with their environment maintained at the relative humidity of 30–40%. All the experimental animal procedures conformed to the guidelines of the Beijing University of Chinese Medicine Animal Care and Use Committee. All experimental protocols were reviewed and approved by the Animal Experimentation Ethics Committee at the Beijing University of Chinese Medicine (BUCM-4-2019042503-2098) and the Ethics Committee of the Third affiliated Hospital of Beijing University of Chinese Medicine (20190228-1.0). All the work tried to minimize animal suffering.
TS AND COMORBID ANXIETY MODEL:
After 1 week of adaptive feeding, a total of 48 rats were randomly divided into the control group (n=12) and a TS model group (n=36). The model was induced by injecting them with IDPN (250 mg/kg−1) once daily, and the control group was intraperitoneally injected with an equal volume of normal saline for 7 consecutive days. Meanwhile, rats in the TS model group were provided water only between 9: 00–9: 10 in the morning and 21: 00–21: 10 in the evening. There was no longer a water supply given during the remainder of the time; the control group was allowed to drink normally. After 7 days, a stress experiment was performed on the TS model group, and the empty bottle was stimulated to induce emotional stress during the regular feeding time. The stimulation was irregular and maintained once a day for 2 weeks (Table 1). The control group was allowed to drink normally. Based on the examination of stereotypy behavior and the elevated plus maze test results, the TS model group was randomly divided into the TS and comorbid anxiety model group (Model group), fluoxetine hydrochloride group (Flx group), and JPZDD group, with 12 rats in each group (Table 1).
DRUG PREPARATION AND ADMINISTRATION:
Drug administration was given immediately after 3 weeks of empty bottle stimulation. JPZDD granules were provided by the Pharmacy Department of the Third Affiliated Hospital of Beijing University of Chinese Medicine identified by Director Xiang-wen Kong. JPZDD included 10 different Chinese medicinal herbs (Table 2). The granules were dissolved in 100 mL of distilled water, well-mixed, and the solution was stored at 4°C before further use. The rats in the JPZDD group received gavage with the solution of JPZDD 16 g/kg−1/d−1. The Flx group was administered a fluoxetine hydrochloride suspension, 4.2 mg/kg−1/d−1 (0943A; Patheon France, Jiangsu, People’s Republic of China), which was equivalent to 4.2 times the clinical dose for children. The control group and model group were administered the same amount of saline. The rats in the 4 groups were gavaged by the intragastric administration of 10 mL/kg−1/d−1 once a day for 4 weeks. The rats were then anesthetized and sacrificed for further study.
BODY WEIGHT:
The body weight was measured at 8: 30 a.m. in the morning after the administration for 1, 2, and 4 weeks.
STEREOTYPY RECORDING:
The rats were placed in the observation cage and adapted for 5 minutes. The double-blind method was used to score the stereotypy behavior of the rats. Two trained observers conducted the test at 1 hour after administration for 1, 2, and 4 weeks. They were familiar with the observation methods, but were blind to the group conditions. Each rat was observed for 2 minutes before the total score was recorded [15]. The scoring criteria were as follows: 0 point, no stereotypes; 1 point, rotation behavior; 2 points, excessive movement of the head and neck; 3 points, excessive movement of the head and neck, plus rotation behavior; 4 points, head to side, continuous sniffing, biting, head bobbing, and repetitive body raising/wall climbing in place [15].
ELEVATED PLUS MAZE TEST:
The elevated plus maze (EPM) was produced by an internationally accepted method and was manufactured by Huaibei Zhenghua Biological Instrument Equipment Co., Ltd. of Anhui Province. EPM is a classic internationally accepted method for measuring anxiety responses. The elevated plus maze is 100 cm above the ground and comprises an open arm (50×10 cm) entry, a closed arm (50×10 cm) entry, and a central area (10×10 cm). The open arm entry and closed arm entry are surrounded by a black plastic wall that is 15 cm high (to prevent the animal from slipping down during the exploration process). Prior to the test, the rats were placed in a plastic box and allowed to freely explore for 3 minutes; the rats were then placed in the middle of the central area before exploration. The test time for each rat was 3 minutes, and the following indicators were recorded: 1) the number of times the rat entered the open arm entry (OE): The duration of activity began when all 4 paws of the rat were in the arm and ended when 1 paw withdrew from the arm; 2) the number of times the rats entered the closed arm entry (CE): The number of times the rats entered any closed arm with all 4 paws. Each rat was tested for 3 minutes; the feces were removed, and the maze was cleaned with a damp cloth and then wiped clean with a dry cloth before testing the next rat. We calculated the ratio of OE and the number of times the rats entered total arm entries (OE+CE)×100%.
HEMATOXYLIN AND EOSIN (HE) STAINING:
At the last of the test, 3 rats from each group were deeply anesthetized with 10% chloral hydrate (4 mL/kg−1), and perfused with chilled physiological saline followed by 4% paraformaldehyde. The brains were removed immediately and fixed in 4% paraformaldehyde for 2 days before paraffin embedding. Three coronal sections of the frontal cortex and striatum were isolated and cut at 4 μm before being mounted on glass slides. The slides were subsequently stained with a modified hematoxylin and eosin-phloxine procedure. The morphological images of the frontal cortex and striatum were captured by a digital camera (BX51T-PHD-J11, Olympus Corporation, Tokyo, Japan).
ENZYME-LINKED IMMUNOSORBENT ASSAY (ELISA):
The rats were anesthetized by intraperitoneal injection with 10% chloral hydrate (4 mL/kg−1) following which 6 mL of blood was taken from the abdominal aorta. The blood was placed in a vacuum blood collection tube at 23±2°C and centrifuged at 2500 rpm for 15 minutes; the serum was then separated and spit into small tubes to store at −20°C. The level of 5-HT in the serum was determined by enzyme-linked immunosorbent assay (ELISA), and the experimental procedures were performed according to the kit instructions (20171010. 60087R, RIGORBIO, Beijing, China). Then 100 μL each of the standard sample and the samples were added (undiluted, 2-well repeat), and reacted at 37°C for 1 hour. We discarded the liquid in the well plate and filled each well with the washing solution, which was then kept still for 30 seconds and discarded. This procedure was repeated 5 times. After drying, we added 50 μL of horseradish peroxidase-labeled avidin working solution, which was allowed to react at 37°C for 1 hour. The solution was soaked for 30 seconds, washed 5 times, and kept dry; 50 μL of color developer A was added to each well; then 50 μL of developer B was mixed in and allowed to develop at 37°C for 10 minutes in the dark. Lastly, 50 μL of stop solution was added to each well to stop the reaction (when the blue color turns yellow). The blank holes were set to zero, and the absorbance (OD value) of each well was measured. The measurement was carried out within 10 minutes after the addition of the stop solution. The regression equation of the standard curve was calculated according to the concentration and the OD value.
HIGH PERFORMANCE LIQUID CHROMATOGRAPHY (HPLC):
The cortex and striatum of both sides were dissected and frozen at −80°C. The striatum of the left side (n=8 or 9, per group) were weighed and then homogenized at 400 μL of ice-cold lysis buffer. Then the homogenate was centrifuged at 15000 rpm for 10 minutes at 4°C and filtered through a 0.22 μm filter (Costar, Spin-X) and then centrifuged at 7000 rpm. Standard solution or the filtrate obtained from brain homogenate (20 μL) was added. Conditions of chromatography were as follows: The chromatographic column was ESA MD-150 (3.2×150 mm, 5 μM). The mobile phase was composed of 90 mM disodium hydrogen phosphate, 50 mM citric acid, 1.7 mM Sodium 1-octane sulfonate, 50 μM of 0.1 M EDTA, and 10% acetonitrile (pH 6.70). The column temperature was maintained at an ambient temperature of 30°C. The flow rate was set to 0.6 mL/minute [16].
WESTERN BLOT:
Protein was extracted from the right cortex and striatum of each rat (n=6), and the protein concentration was determined using BSA as the standard. Samples containing 40 μg of protein were added into the 10% polyacrylamide slab gel. After the protein was transferred to nitrocellulose membranes, the membranes were blocked by 5% skimmed milk powder in TBS-T for 60 minutes and then incubated with the primary antibody (5HT2C: LS-B11334, 1: 1000, LSBIO, Seattle, WA, USA) and GAPDH (ab8245, 1: 10000, Abcam, Cambridge, MA, USA) at 23±2°C overnight. After washing the membrane, the specific binding was detected by goat-anti-rabbit or goat-anti-mouse horseradish peroxidase-conjugated secondary antibodies (5HT2C: ZB-2301, 1: 2000, ZSGB-Bio, Beijing, China; GAPDH: ZB-2305, 1: 5000, ZSGB-Bio, Beijing, China). The staining was visualized using the ECL detection system (SC-2048, Santa Cruz, CA, USA). Images were acquired with an electrophoresis gel imaging analysis system. Band density was measured using Quantity One software (Bio-Rad Laboratories Inc., Hercules, CA, USA).
QUANTITATIVE POLYMERASE CHAIN REACTION (QPCR):
The cortex and striatum on the right side (n=6) of each rat were taken, and total RNA from the cortex and striatum were isolated using TRIzol reagent, according to the manufacturer’s protocol (15596018, Invitrogen, Carlsbad, CA, USA). The quality and quantity of the isolated RNA were tested using agarose gel electrophoresis and a spectrophotometer (260/280 ratio; Nanodrop 2000; Thermo Fisher Scientific, Waltham, MA, USA). The cDNA was synthesized using a reverse transcription kit under the following conditions. The basic protocol for quantitative polymerase chain reaction (qPCR) comprised an initial denaturation at 94°C for 10 minutes, followed by 45 cycles of amplification. For cDNA amplification, the cycles consisted of 94°C for 15 seconds and 60°C for 60 seconds. The final elongation step was conducted at 72°C for 10 minutes. The gene primers were as follows:
The PCR experiments were carried out using a qPCR machine (ABI7500, Applied Biosystems, Waltham, MA, USA), and transcript levels was determined using the 2−ΔΔCt method.
STATISTICAL ANALYSIS:
All statistical data were expressed as the mean±standard deviation and analyses performed using IBM SPSS 20.0 statistical software (IBM, Armonk, NY, USA). Statistical analysis was performed using either one-way analysis of variance ANOVA (equal variance) or a Welch’s ANOVA (unequal variance) test. Differences were considered to be statistically significant for
Results
EFFECT OF JPZDD ON BODY WEIGHT:
The body weight was measured at 1, 2, and 4 weeks (Figure 1). No significant difference could be seen between the groups at the first and second week after the drug administration. At week 4, the body weights of the model group (140.33±13.64 g), the JPZDD group (152.00±6.00 g), and the Flx group (152.17±15.94 g) were significantly lower than that of the control group (164.83±8.24 g) (P<0.01). The weight of the Flx group and the JPZDD group were significantly higher than that of the model group (P<0.05 for both).
EFFECT OF JPZDD ON STEREOTYPY BEHAVIOR:
Stereotypy behavior tests were performed at 1, 2, and 4 weeks (Figure 2). At the end of week 1, no significant difference could be seen between the groups. At 2 weeks, the stereotypy score of the JPZDD group (1.67±0.89) was lower than that of the model group (2.92±1.17) (P<0.01) and the Flx group (2.58±1.08) (P<0.05). At 4 weeks, the stereotypy behavior in the JPZDD group (1.67±1.07) was still less than that of the model group (2.83±0.94) (P<0.01), but no statistical difference could be seen when compared with the Flx group. These results suggest that JPZDD could alleviate the stereotypy behavior of TS and comorbid anxiety rats.
EFFECT OF JPZDD ON ELEVATED PLUS MAZE SCORE:
The elevated plus maze test was performed at 1, 2, and 4 weeks, and statistically significant differences between the groups could be seen at each time point (P<0.05, P<0.01, P<0.05, respectively; Figure 3). At the end of week 1, the ratio of OT/TT decreased significantly in the model group (29.83±8.03) compared with the control group (63.75±7.32) (P<0.01). The ratio of OT/TT increased more higher in the JPZDD group (57.58±9.20) than the model group (P<0.05). At 2 weeks, when compared with the control group (61.0±9.39), the ratio of OT/TT decreased significantly in the model group (26.83±4.81) (P<0.01), and the ratio of OT/TT in the JPZDD group (53.17±29.10) was higher than the model group (P<0.05). At 4 weeks, the model group (32.67±8.41) still showed a downward trend when compared with the control group (55.5±7.02) (P<0.05). The ratio of OT/TT was higher in the JPZDD group (55.67±8.65) than the model and Flx groups (32.33±3.86) (P<0.05).
EFFECT OF JPZDD ON MORPHOLOGIC CHANGES:
The cortical cells in the control group were arranged neatly, and the cell structure could be seen clearly. The cortical cells in the model group were arranged loosely with obvious cell destruction and nuclear pyknosis. The cytoplasmic and cytosolic boundaries were not clear and there was vacuolization. The cortical cells in the Flx group showed some improvement compared with the model group. The nucleus of the cortical cells showed slight pyknosis and the cells were arranged neatly. The neural number in the JPZDD group increased, and the neuronal structure was normal.
The striatal cells in the control group were closely linked. The nucleus was large and round. The distribution of chromatin was evenly distributed, whereas in the model group, the cells were disorganized. The division of cytoplasm and the nucleus was unclear, and nuclear pyknosis and vacuolization could be seen. In the Flx and JPZDD groups, the nucleus of the striatal cells showed slight pyknosis, the cells were arranged neatly, and the structures of the neurons were gradually normal (Figure 4).
EFFECT OF JPZDD ON 5-HT LEVELS:
The levels of 5-HT were examined by high performance liquid chromatography (HPLC) and ELISA. Compared with the control group (0.03±0.01 ng/mg), the level of 5-HT in the striatum of the model group (0.01±0.00 ng/mg) decreased significantly (P<0.01). After the treatment, the 5-HT levels in the JPZDD (0.02±0.00 ng/mg) and Flx groups (0.02±0.01 ng/mg) increased. Statistically significant difference between the JPZDD group and the model group could be seen (P<0.01). And the same difference could be seen between the Flx group and the model group (P<0.01). But there was no statistical difference between the JPZDD group and Flx group. The serum level of 5-HT in the model group (79.27±3.71 ng/mL) decreased significantly compared with the control group (121.82±5.28 ng/mL) (P<0.01). After the treatment, the levels of 5-HT in the JPZDD (96.0±4.09 ng/mL) and Flx groups (88.73±4.09 ng/mL) increased. The 5-HT levels in the JPZDD group was higher than the model group (P<0.01) (Figure 5).
EFFECT OF JPZDD ON 5-HT2C EXPRESSION:
Compared with the control group (1.00±0.01), the expression of 5-HT2C in the cortex of the model group (0.50±0.27) showed a downward trend (P<0.01). The expression in the cortex of the Flx (0.49±0.20) and JPZDD group (0.43±0.13) was also lower than that of the control group (P < 0.01), but there was no statistical difference between the treatment groups and the model group. The expression of 5-HT2C in the striatum also showed a decreasing trend, but there was no statistical difference among the groups (Figure 6A).
EFFECT OF JPZDD ON 5-HT2C MRNA TRANSCRIPT EXPRESSION:
Compared with the control group (1.00±0.06, 1.00±0.07), the expression of mRNA in the cortex and striatum of the model group (1.16±0.07, 1.58±0.21) showed an upward trend (P<0.01, P<0.01, respectively). After 4 weeks of treatment, the expression of mRNA in the striatum decreased, and the difference between the JPZDD group (1.02±0.16) and the model group was statistically significant (P<0.01). Compared with the Flx group(1.49±0.11), the JPZDD group showed a significant decreasing trend (P<0.01) (Figure 6B).
Discussion
TS is a childhood onset disorder characterized by motor and vocal tics and is associated with multiple coexisting neuropsychological difficulties including anxiety and ADHD [17]. The complexity of TS, which results from a myriad of symptoms and the fact that the pathophysiology leading to the disorder is still largely unknown, has resulted in high variability in studies examining the mechanisms and behavioral symptoms in animal models of TS [18]. In the present study, a TS and comorbid anxiety model was established by IDPN intraperitoneal injection combined with uncertain empty bottle stimulation. TS was induced by IDPN intraperitoneal injection, and uncertain empty bottle stimulation contributed to the development of anxiety. Previous studies had verified that this model had better surface validity and structural validity. The face validity was verified by stereotypy score and elevated plus maze, the construct validity was verified by the content of 5-HT using HPLC. Its neurotransmitter changes were similar to those of children with TS and comorbid anxiety, and that it could be used as an animal model for studying TS and comorbid anxiety [14].
According to TCM, the pathogenesis of TS involves spleen deficiency and live fire. The comorbidity of anxiety is related with heart-shen. JPZDD is made from Xie-qing-wan (XQW) and Liu-jun-zi decoction (LJZD). XQW can suppress the liver fire and LJZD can strengthen the spleen, so as to calm heart-shen. There are 10 Chinese herbs in JPZDD [19]; Gentiana scabra Bge and Pseudostellaria heterophylla Pax are the chief drugs, whereas Poria cocos Wolf, Atractylodes macrocephala Koidz, Citrus reticulata Blanco, and Pinellii ternata Breit are adjuvant herbs. Others are assistant herbs. Previous animal experiments have confirmed that JPZDD can regulate the levels of monoamines and amino acid transmitters and their related receptors and transporters in the brain [19,20] and improve the symptoms of TS comorbid mood disorders [21]. However, it is still not clear the therapeutic mechanism of JPZDD.
In this study, behavioral test results showed that at the fourth week of treatment, the body weight of the model group was significantly lower than that of the control group, while the weight of rats in the Flx group and the JPZDD group were higher than that of the model group. The results of the stereotypy exercise test showed that the stereotypy exercise score of the JPZDD group was lower than that of the model group at the second week of treatment, indicating that JPZZD had a good effect on improving tic symptoms by alleviating stereotypy behavior. This study also included elevated plus maze testing on the rats. The results showed that after 1 week of administration, the OT/TT ratio was lower in the model group than the control group (
It is generally agreed upon that abnormalities in the CSTC pathways, with an imbalance of inhibitory and excitatory pathways, have an important effect on the pathogenesis of TS [22]. TS was associated with a reduced number of neurons in the striatum. Previous studies indicated that a striatal fast-spiking neuron deficit can produce stress-triggered repetitive movements and anxiety [23]. We identified morphologic changes in both the striatum and the cortex of the TS and comorbid anxiety rats in our study. The cells in the model group were disorganized, and nuclear pyknosis and vacuolization could be seen. After the treatments, the nucleus of both the striatal and cortex cells showed slight pyknosis, the structure of the neurons became gradually normal, and the cells were arranged neatly. These results indicated that JPZDD could reverse the morphologic changes, but the exact changes were still not clear. New technologies and further studies will make efforts to elucidate these findings.
The pathogenesis and development of TS and anxiety are closely related to the basal ganglia DA and 5-HT neurotransmitter system [24]. The general consensus is that the excitability of the DA system is increased, while the 5-HT transmitter system is absolutely or relatively insufficient. A change of one neurotransmitter often influences the function of other interconnected transmitters significantly, so the change of the neurotransmitters may be a dynamic indicator in the pathophysiology of tics and anxiety. 5-HT plays a general inhibitory and stabilizing role in the high-level parts of the brain and mental activity [25]. Metabolism of 5-HT in the brain directly affects or is related to human mental activity. A large number of studies have shown that 5-HT metabolic abnormalities play important role in the pathophysiological mechanism of TS [26], anxiety disorders [27], ADHD [28], OCD [29,30] and other mental behavior disorders. Fluoxetine, a selective serotonin reuptake inhibitor, as the first-line psychological treatment to counter TS and comorbid anxiety also supports the association of 5-HT dysfunction with TS and comorbid anxiety [11]. In addition to being effective for the OCD/anxiety, there may be an indirect reduction in tic severity [31], so we choose fluoxetine as the control drug. When genetic, developmental, or environmental factors affect, the 5-HT system cannot exert inhibitory control over neuronal activity. So, the symptoms of tics will be more obvious and may also be comorbid with anxiety, OCD, and ADHD. 5-HT neurotransmission is closely related to the pathogenesis of anxiety disorders and tics, mainly due to a decrease in 5-HT transporter expression and an increase in receptor expression [32]. After being released into the synaptic cleft, 5-HT binds with the 5-HT receptors of the postsynaptic membrane so as to exert a biological effect. Currently, 7 serotonin receptor (5-HTR) subtypes have been identified in mammals, and studies have shown that the 5-HT2C receptor is involved in the occurrence of TS [33]. The 5-HT2C receptor is distributed only in the nervous system, including the frontal cortex, hippocampus, hypothalamic nucleus, and striatum. An agonist of the 5-HT receptor exerts an inhibitory effect on the activity of the dopamine system, resulting in decreased dopamine levels, whereas an antagonist of the 5-HT receptor produces an agonistic effect on the activity of the dopamine system, resulting in an increase in dopamine levels. Activating 5-HT2C receptors leads to a decrease in striatal dopamine levels [34], which is the pharmacological basis for its antipsychotic effects.
In this study, we analyzed the mechanism of anti-tic and anti-anxiety effects of JPZDD based on a 5-HT metabolic perspective. The results of this study demonstrated that the 5-HT level both in the striatum and serum of the model group was more lower than control group, indicating that there was a decrease in 5-HT levels in the TS and comorbid anxiety rats. After treatment, the level of 5-HT in the JPZDD group increased significantly in the striatum compared with the model group. This suggests that JPZDD can repair damaged neurons and increase the concentration of 5-HT. This conclusion is consistent with the results reported in most articles, which indicate that the hypofunction of 5-HT neurons is involved in the pathophysiology of TS [32]. In correlation to the receptor, both the cortex and striatum were observed. The protein expression levels of the 5-HT2C receptor demonstrated that the expression of 5-HT2C in the cortex of both the model group and 2 treatment groups exhibited a decreasing trend when compared with the control group (
In summary, the aforementioned results indicate that an IDPN intraperitoneal injection combined with an uncertain empty bottle stimulation model can simulate the tics and anxiety behavior of TS and comorbid anxiety rats. There was a feedback regulation of the 5-HT system in the TS and comorbid anxiety rats, but its activity was relatively insufficient. JPZDD could improve the symptoms of tic and anxiety, and its mechanism involves reducing the expression of 5-HT2C mRNA in the striatum and increasing the concentration of 5-HT so as to exert its neuroinhibitory effects. The results of our data suggested that JPZDD was a potential therapeutic intervention for TS and comorbid anxiety.
Overall, this study tried to explore the effects of JPZDD on TS and comorbid anxiety. We mainly identified its effects on the 5-HT system. Both the content of 5-HT and its receptor in the cortex and striatum were observed. Understanding these regulatory mechanisms may facilitate the development of therapies for TS and comorbid anxiety treatment. But the present study still had some limitations. The exact mechanism of each Chinese herb in JPZDD should be validated by fingerprint of traditional Chinese medicine, and other studies on the content of neurotransmitters in the brain are needed to provide molecular biological evidence for the mechanism of JPZDD. Other behavioral tests, such as thigmotaxia as a test for anxiolytic activity, should be done. The contributions of other receptors or transporters of 5-HT and other signaling circuits involved in the neuroprotective effects of JPZDD should be the subject of further studies.
Conclusions
5-HT neurotransmission and its receptors are closely related to the pathogenesis of TS and comorbid anxiety disorder within the CSTC brain circuit. JPZDD alleviated both the tic and anxiety symptoms of rats with TS and comorbid anxiety, and the mechanism may be via reducing the expression of 5-HT2C mRNA in the striatum, increasing the concentration of 5-HT, and enhancing the activity of the 5-HT system, which in turn exerts neuro-inhibition.
Figures
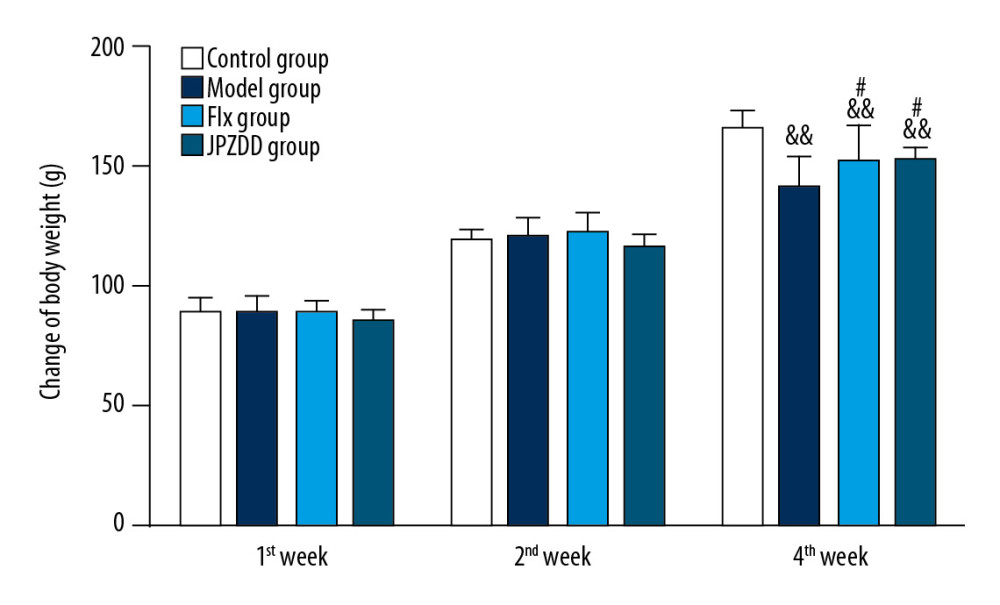 Figure 1. Effect of treatment on the body weight of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=1.26, P=0.24; 2 weeks: F=1.54, P=0.22; 4 weeks: F=10.76, P=0.00) && P<0.01 versus the control group; # P<0.05 versus the model group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 1. Effect of treatment on the body weight of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=1.26, P=0.24; 2 weeks: F=1.54, P=0.22; 4 weeks: F=10.76, P=0.00) && P<0.01 versus the control group; # P<0.05 versus the model group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group. 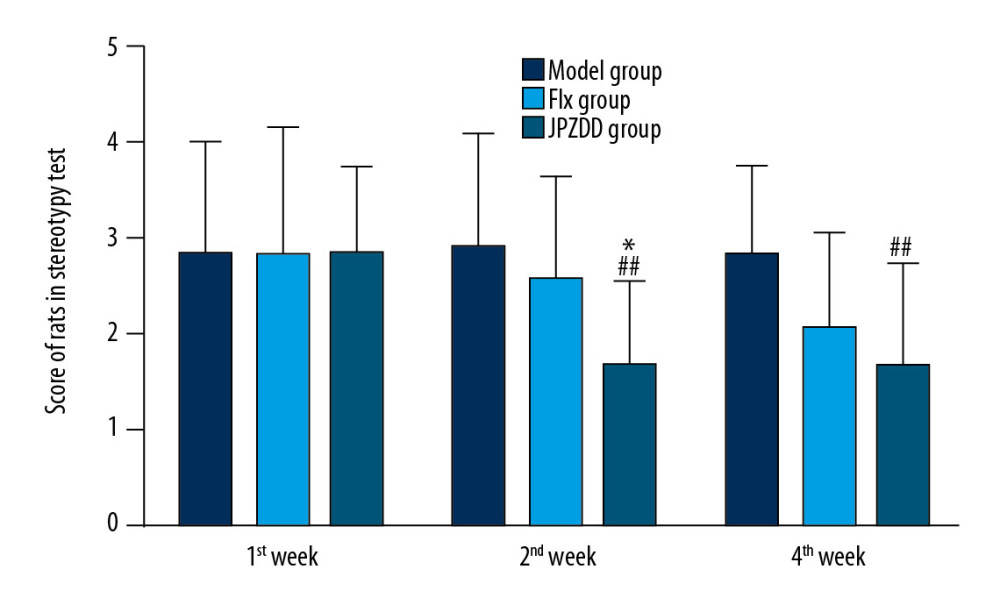 Figure 2. Effect of treatment on the stereotypy behavior score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=0.00, P=1.00; 2 weeks: F=4.55, P=0.02; 4 weeks: F=4.16, P=0.02); ## P<0.01 vs. the model group; * P<0.05 vs. the Flx group; Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 2. Effect of treatment on the stereotypy behavior score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=0.00, P=1.00; 2 weeks: F=4.55, P=0.02; 4 weeks: F=4.16, P=0.02); ## P<0.01 vs. the model group; * P<0.05 vs. the Flx group; Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group. 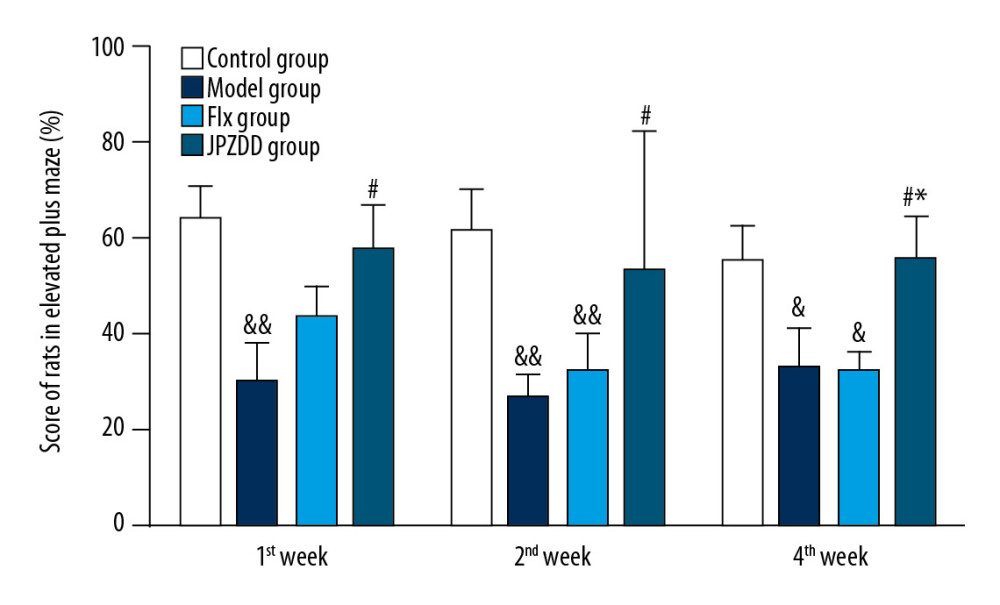 Figure 3. Effect of treatment on the elevated plus maze score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=3.74, P=0.02; 2 weeks: F=4.35, P=0.01; 4 weeks: F=3.39, P=0.03); && P<0.01 versus the control group; & P<0.05 versus the control group; # P<0.05 versus the model group; * P<0.05 versus the Flx group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 3. Effect of treatment on the elevated plus maze score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=3.74, P=0.02; 2 weeks: F=4.35, P=0.01; 4 weeks: F=3.39, P=0.03); && P<0.01 versus the control group; & P<0.05 versus the control group; # P<0.05 versus the model group; * P<0.05 versus the Flx group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group. 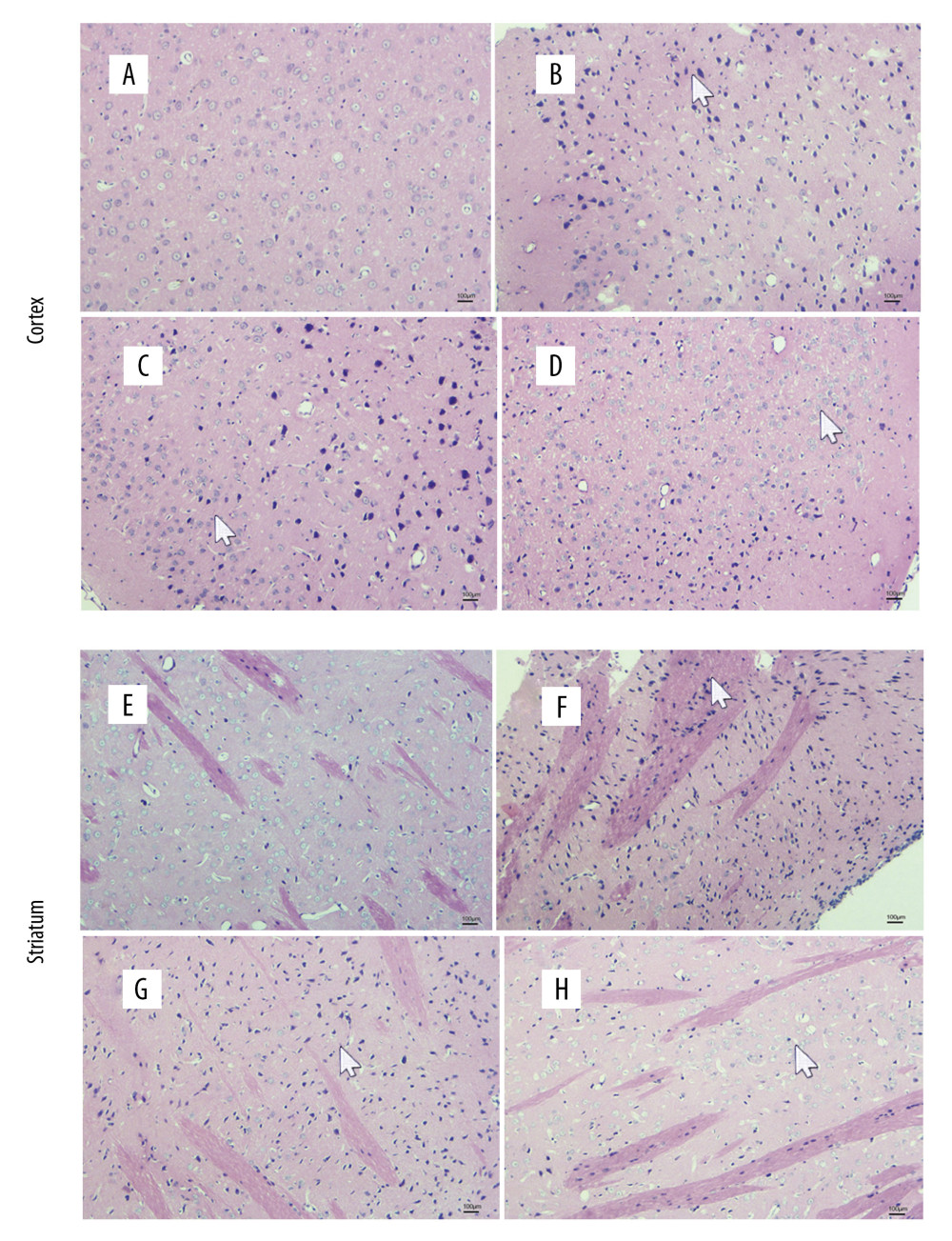 Figure 4. Alternations of morphologic changes in the cortical and striatal sections stained with hematoxylin and eosin. The sections were viewed at 100× magnification. Scale bar=100 μm (n=3 per group). (A) Cortex changes in the control group. (B) Cortex changes in the model group. (C) Cortex changes in the Flx group. (D) Cortex changes in the JPZDD group. (E) Striatum changes in the control group. (F) Striatum changes in the model group. (G) Striatum changes in the Flx group. (H) Striatum changes in the JPZDD group. The white arrow points to the morphologic changes of each group. Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride-treated group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 4. Alternations of morphologic changes in the cortical and striatal sections stained with hematoxylin and eosin. The sections were viewed at 100× magnification. Scale bar=100 μm (n=3 per group). (A) Cortex changes in the control group. (B) Cortex changes in the model group. (C) Cortex changes in the Flx group. (D) Cortex changes in the JPZDD group. (E) Striatum changes in the control group. (F) Striatum changes in the model group. (G) Striatum changes in the Flx group. (H) Striatum changes in the JPZDD group. The white arrow points to the morphologic changes of each group. Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride-treated group; JPZDD group, Jian-pi-zhi-dong decoction group. 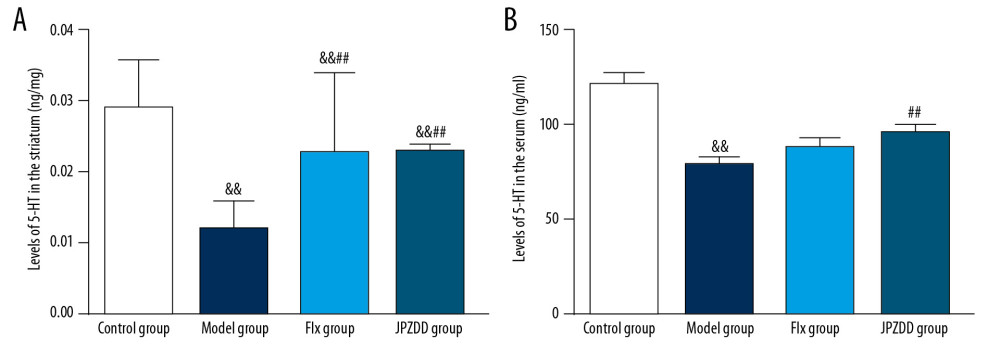 Figure 5. Effect of JPZDD on 5-HT levels. Data are presented as mean±standard deviation. (A) n=9 for the control group, n=8 for the other groups (F=15.52, P<0.01) (B) n=10 for the control group, Flx group, and JPZDD group; n=12 for the model group (F=18.57, P<0.01). && P<0.01 versus the control group; ## P<0.01 versus the model group; Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 5. Effect of JPZDD on 5-HT levels. Data are presented as mean±standard deviation. (A) n=9 for the control group, n=8 for the other groups (F=15.52, P<0.01) (B) n=10 for the control group, Flx group, and JPZDD group; n=12 for the model group (F=18.57, P<0.01). && P<0.01 versus the control group; ## P<0.01 versus the model group; Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group. 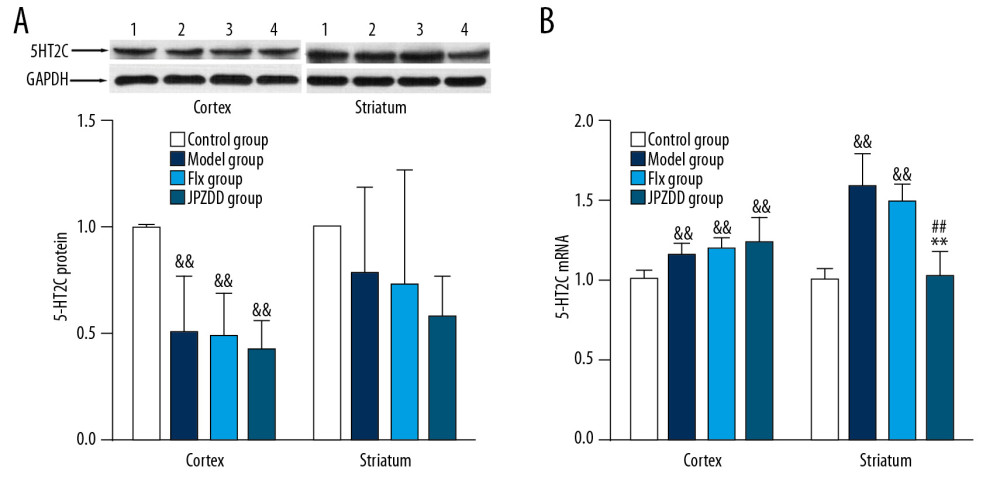 Figure 6. Effects of treatment on the expression levels of 5-HT2C in the cortex and striatum of Tourette syndrome (TS) and comorbid anxiety rats. Western blot and quantitative polymerase chain reaction were performed to investigate changes in the expression of 5-HT2C and the effects of JPZDD on those levels. && P<0.01 versus the control group; ## P<0.01 versus the model group; ** P<0.01 versus Flx group. (A) 5-HT2C protein expression (n=3 per group, F=20.15, P=0.01; F=3.76, P=0.14). 1=control group; 2=model group; 3=Flx group; 4=JPZDD group. (B) 5-HT2C mRNA expression (n=6 per group, F=6.54, P=0.00; F=34.14, P=0.00). Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.
Figure 6. Effects of treatment on the expression levels of 5-HT2C in the cortex and striatum of Tourette syndrome (TS) and comorbid anxiety rats. Western blot and quantitative polymerase chain reaction were performed to investigate changes in the expression of 5-HT2C and the effects of JPZDD on those levels. && P<0.01 versus the control group; ## P<0.01 versus the model group; ** P<0.01 versus Flx group. (A) 5-HT2C protein expression (n=3 per group, F=20.15, P=0.01; F=3.76, P=0.14). 1=control group; 2=model group; 3=Flx group; 4=JPZDD group. (B) 5-HT2C mRNA expression (n=6 per group, F=6.54, P=0.00; F=34.14, P=0.00). Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group. 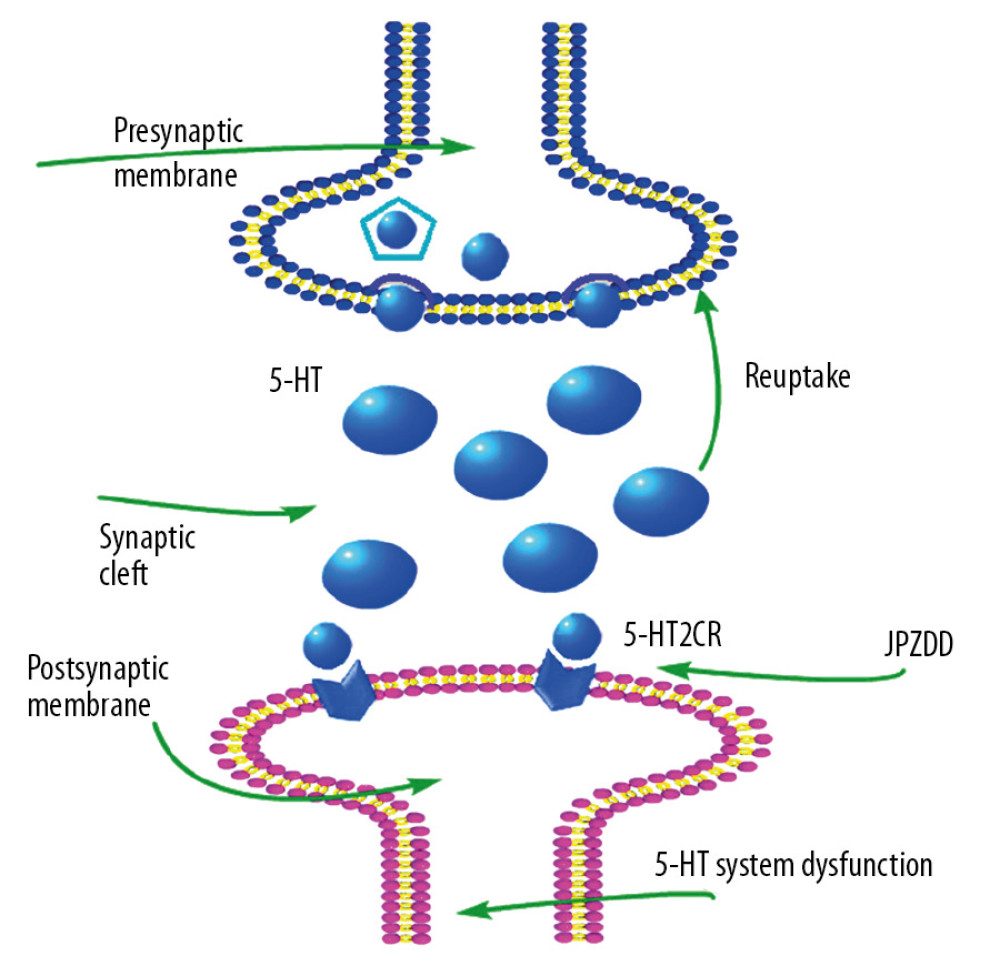 Figure 7. Schematic representation of JPZDD effects on 5-HT system. JPZDD exerts an anti-tic and anti-anxiety effect on the 5-HT system in a rat model of Tourette syndrome (TS) and comorbid anxiety mainly by inhibiting the expression of 5-HT2C mRNA in the striatum, improved the relative deficiency of 5-HT, and further enhanced the excitability of 5-HT system. 5-HT, serotonin; 5-HT2CR, 5-HT type 2C receptor; JPZDD, Jian-pi-zhi-dong decoction.
Figure 7. Schematic representation of JPZDD effects on 5-HT system. JPZDD exerts an anti-tic and anti-anxiety effect on the 5-HT system in a rat model of Tourette syndrome (TS) and comorbid anxiety mainly by inhibiting the expression of 5-HT2C mRNA in the striatum, improved the relative deficiency of 5-HT, and further enhanced the excitability of 5-HT system. 5-HT, serotonin; 5-HT2CR, 5-HT type 2C receptor; JPZDD, Jian-pi-zhi-dong decoction. References
1. American Psychiatric Association: Diagnostic and statistical manual of mental disorders, 2013, Washington, DC, American Psychiatric Association
2. Ganos C, Martino D, Tics and Tourette syndrome: Neurol Clinics, 2015; 33(1); 115-36
3. Gillberg C, Billstedt E, Autism and Asperger syndrome: Coexistence with other clinical disorders: Acta Psychiat Scand, 2000; 102(5); 321-30
4. Marwitz L, Pringsheim T, Clinical utility of screening for anxiety and depression in children with Tourette syndrome: J Can Acad Child Adolesc Psychiatry, 2018; 27(1); 15-21
5. Hirschtritt ME, Lee PC, Pauls DL, Comorbid psychiatric disorders in Tourette syndrome: JAMA Psychiat, 2015; 72(4); 325-33
6. Coffey BJ, Biederman J, Smoller JW, Anxiety disorders and tic severity in juveniles with Tourette’s disorder: J Am Acad Child Adolesc Psychiatry, 2000; 39(5); 562-68
7. Chen J, Leong PK, Leung H, A Chinese herbal formulation, Xiao-Er-An-Shen decoction, attenuates Tourette syndrome, possibly by reversing abnormal changes in neurotransmitter levels and enhancing antioxidant status in mouse brain: Front Pharmacol, 2019; 10; 812
8. Robertson MM, Eapen V, Singer HS, Gilles de la Tourette syndrome: Nat Rev Dis Primers, 2017; 3; 16097
9. Ji XY, Wu M, Zhang X, Dopamine level in the stratium exhibits circadian rhythms in the rat model of Tourette syndrome: Int J Clin Exp Med, 2016; 9(6); 9015-20
10. Sant’Ana AB, Vilela-Costa HH, Vicente MA, Role of 5-HT2C receptors of the dorsal hippocampus in the modulation of anxiety- and panic-related defensive responses in rats: Neuropharmacology, 2019; 148; 311-19
11. Rickards H, Cavanna AE, Worrall R, Treatment practices in Tourette syndrome: the European perspective: Eur J Paediatr Neurol, 2012; 16(4); 361-64
12. Zhang W, Wang SM, Yu WJ, Application of TCM theory of brain spirit and Wuzang spirit in treatment of the comorbidity of Tourette syndrome: J Hunan Univ Chin Med, 2019; 29(2); 203-6
13. Yu WJ, Zhang W, Shi XW, Effects of Jianpizhidong decoction on expression of serum BDNF in Tourette syndrome children: Chin Arch Tradit Chin Med, 2019; 37(2); 79-82
14. Zhang W, Cui X, Yu WJ, Assessment of a rat model for comorbidity of Tourette syndrome and anxiety with empty water bottle stimulation plus iminodipropionitrile injection: Chinese J Behav Med Brain Sci, 2017; 26(9); 775-81
15. Khan HA, Alhomida AS, Arif IA, Neurovestibular toxicities of acrylonitrile and iminodipropionitrile in rats: A comparative evaluation of putative mechanisms and target sites: Toxicol Sci, 2009; 109(1); 124-31
16. Zhang F, Li AY, Dual ameliorative effects of Ningdong granule on dopamine in rat models of Tourette’s syndrome: Sci Rep, 2015; 5; 7731
17. Singer HS, Tics and Tourette syndrome: Continuum (Minneap Minn), 2019; 25(4); 936-58
18. Dorin Y, Michal I, Izhar BG, Animal models of Tourette syndrome – from proliferation to standardization: Front Neurosci, 2016; 10; 132
19. Zhang W, Wei L, Yu W, Effect of Jian-Pi-Zhi-Dong decoction on striatal glutamate and γ-aminobutyric acid levels detected using microdialysis in a rat model of Tourette syndrome: Neuropsych Dis Treat, 2016; 12; 1233-42
20. Yu WJ, Shi XW, Cui X, Jian-pi-zhi-dong-decoction regulates the expression of glutamate transporters to attenuate glutamate excitotoxicity and exerts anti-tics effects in Tourette syndrome model rats: Neuropsych Dis Treat, 2018; 14; 3381-92
21. Cui X, Yu WJ, Zhang W, Effects of Jian-pi-zhi-dong decoction on hypothalamic pituitary adrenal axis of children with Tourette syndrome: Mod Chinese Clin Med, 2016; 23(6); 17-20
22. Mahone EM, Puts NA, Edden RAE, GABA and glutamate in children with Tourette syndrome: A 1H MR spectroscopy study at 7 T: Psychiat Res Neuroimaging, 2018; 273; 46-53
23. Xu M, Li L, Pittenger C, Ablation of fast-spiking interneurons in the dorsal striatum, recapitulating abnormalities seen post-mortem in Tourette syndrome, produces anxiety and elevated grooming: Neuroscience, 2016; 324; 321-29
24. Bacque CJ, Bharatiya R, Barriere G, Serotonin in animal cognition and behavior: Int J Mol Sci, 2020; 21(5); 1649
25. Temel Y, High frequency stimulation of the subthalamic nucleus: Linking mood and motor effects at the level of the basal ganglia and 5-HT system: Brain Simul, 2019; 12(2); 412
26. Quezada J, Coffman KA, Current approaches and new developments in the pharmacological management of Tourette syndrome: CNS Drugs, 2018; 32(1); 33-45
27. Marcinkiewcz CA, Mazzone CM, D’Agostino G, Serotonin engages an anxiety and fear-promoting circuit in the extended amygdala: Nature, 2016; 537; 97-101
28. Stewart A, Davis GL, Gresch PJ, Serotonin transporter inhibition and 5-HT2C receptor activation drive loss of cocaine-induced locomotor activation in DAT Val559 mice: Neuropsychopharmacol, 2019; 44; 994-1006
29. Müller-Vahl KR, Szejko N, Wilke F, Serotonin transporter binding is increased in Tourette syndrome with obsessive compulsive disorder: Scie Rep, 2019; 9; 972
30. Chagraoui A, Thibaut F, Skiba M, 5-HT2C receptors in psychiatric disorders: A review: Prog Neuropsychopharmacol Biol Psychiatry, 2016; 66; 120-35
31. Efron D, Dale RC, Tics and Tourette syndrome: J Paediatr Child Health, 2018; 54(10); 1148-53
32. Nomura Y, Tourette syndrome: Clinical features and pathophysiology: Brain Nerve, 2017; 69(12); 1373-85
33. Dehning S, Müller N, Matz J, A genetic variant of HTR2C may play a role in the manifestation of Tourette syndrome: Psychiat Genet, 2010; 20(1); 35-38
34. Burke MV, Christine N, Sonneborn AJ, Striatal serotonin 2C receptors decrease nigrostriatal dopamine release by increasing GABA-A receptor tone in the substantia nigra: J Neurochem, 2014; 131(4); 432-43
35. Pandey S, Dash D, Progress in pharmacological and surgical management of Tourette syndrome and other chronic tic disorders: Neurologist, 2019; 24(3); 93-108
36. Sant’Ana AB, Vilela-Costa HH, Vicente MA, Role of 5-HT2C receptors of the dorsal hippocampus in the modulation of anxiety- and panic-related defensive responses in rats: Neuropharmacol, 2019; 148; 311-19
37. Demireva EY, Deepika S, Emanuela M, 5-HT2 Creceptor blockade reverses SSRI-associated basal ganglia dysfunction and potentiates therapeutic efficacy: Mol Psychiatr, 2018 [Online ahead of print]
Figures
Figure 1. Effect of treatment on the body weight of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=1.26, P=0.24; 2 weeks: F=1.54, P=0.22; 4 weeks: F=10.76, P=0.00) && P<0.01 versus the control group; # P<0.05 versus the model group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 2. Effect of treatment on the stereotypy behavior score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=0.00, P=1.00; 2 weeks: F=4.55, P=0.02; 4 weeks: F=4.16, P=0.02); ## P<0.01 vs. the model group; * P<0.05 vs. the Flx group; Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 3. Effect of treatment on the elevated plus maze score of Tourette syndrome (TS) and comorbid anxiety rats. Data are presented as mean±standard deviation. (n=12; 1 week: F=3.74, P=0.02; 2 weeks: F=4.35, P=0.01; 4 weeks: F=3.39, P=0.03); && P<0.01 versus the control group; & P<0.05 versus the control group; # P<0.05 versus the model group; * P<0.05 versus the Flx group. Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 4. Alternations of morphologic changes in the cortical and striatal sections stained with hematoxylin and eosin. The sections were viewed at 100× magnification. Scale bar=100 μm (n=3 per group). (A) Cortex changes in the control group. (B) Cortex changes in the model group. (C) Cortex changes in the Flx group. (D) Cortex changes in the JPZDD group. (E) Striatum changes in the control group. (F) Striatum changes in the model group. (G) Striatum changes in the Flx group. (H) Striatum changes in the JPZDD group. The white arrow points to the morphologic changes of each group. Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride-treated group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 5. Effect of JPZDD on 5-HT levels. Data are presented as mean±standard deviation. (A) n=9 for the control group, n=8 for the other groups (F=15.52, P<0.01) (B) n=10 for the control group, Flx group, and JPZDD group; n=12 for the model group (F=18.57, P<0.01). && P<0.01 versus the control group; ## P<0.01 versus the model group; Model group, Tourette syndrome (TS) and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 6. Effects of treatment on the expression levels of 5-HT2C in the cortex and striatum of Tourette syndrome (TS) and comorbid anxiety rats. Western blot and quantitative polymerase chain reaction were performed to investigate changes in the expression of 5-HT2C and the effects of JPZDD on those levels. && P<0.01 versus the control group; ## P<0.01 versus the model group; ** P<0.01 versus Flx group. (A) 5-HT2C protein expression (n=3 per group, F=20.15, P=0.01; F=3.76, P=0.14). 1=control group; 2=model group; 3=Flx group; 4=JPZDD group. (B) 5-HT2C mRNA expression (n=6 per group, F=6.54, P=0.00; F=34.14, P=0.00). Model group, TS and comorbid anxiety model group; Flx group, fluoxetine hydrochloride group; JPZDD group, Jian-pi-zhi-dong decoction group.Figure 7. Schematic representation of JPZDD effects on 5-HT system. JPZDD exerts an anti-tic and anti-anxiety effect on the 5-HT system in a rat model of Tourette syndrome (TS) and comorbid anxiety mainly by inhibiting the expression of 5-HT2C mRNA in the striatum, improved the relative deficiency of 5-HT, and further enhanced the excitability of 5-HT system. 5-HT, serotonin; 5-HT2CR, 5-HT type 2C receptor; JPZDD, Jian-pi-zhi-dong decoction. Tables
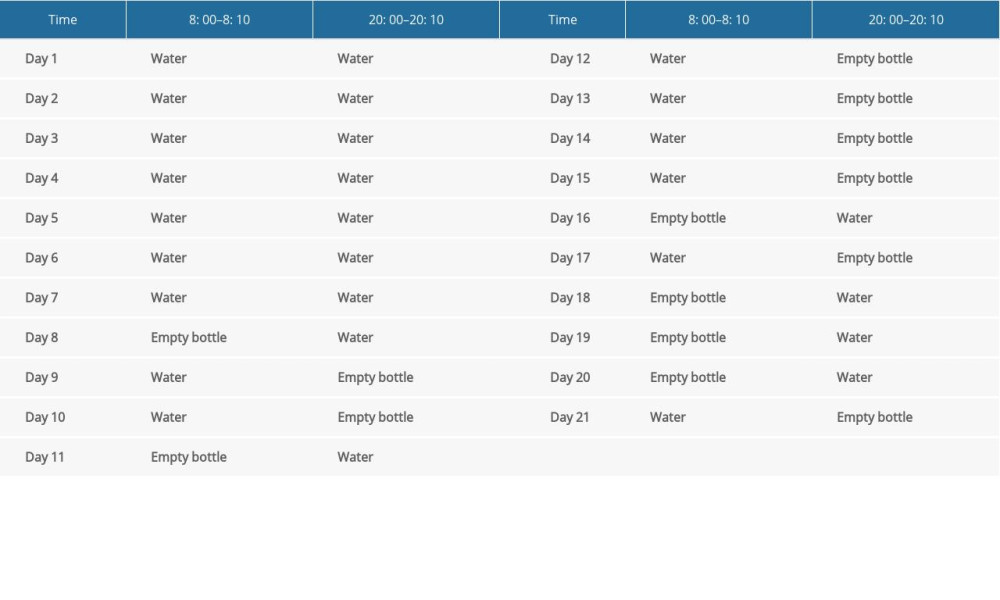 Table 1. A randomized regimen of water feeding in the Tourette syndrome and comorbid anxiety rat model stimulated by an empty bottle.
Table 1. A randomized regimen of water feeding in the Tourette syndrome and comorbid anxiety rat model stimulated by an empty bottle.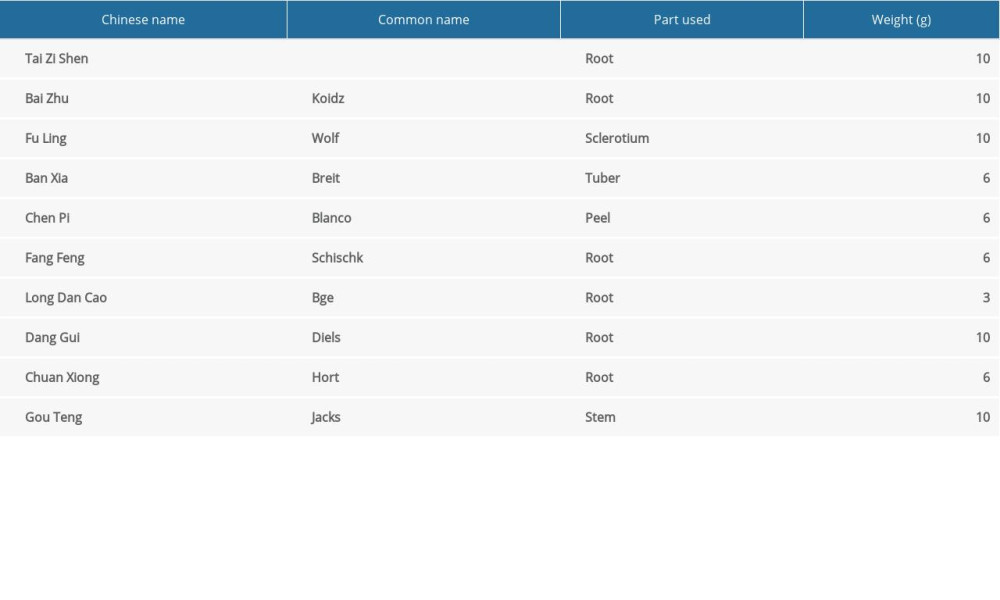 Table 2. The ingredients of Jian-Pi-Zhi-Dong decoction.Table 1. A randomized regimen of water feeding in the Tourette syndrome and comorbid anxiety rat model stimulated by an empty bottle.Table 2. The ingredients of Jian-Pi-Zhi-Dong decoction.
Table 2. The ingredients of Jian-Pi-Zhi-Dong decoction.Table 1. A randomized regimen of water feeding in the Tourette syndrome and comorbid anxiety rat model stimulated by an empty bottle.Table 2. The ingredients of Jian-Pi-Zhi-Dong decoction. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387