04 January 2021: Animal Study
Improves Embryo Survival by Regulating Th1/Th2 Balance in a Mouse Model of Recurrent Spontaneous Abortion
Jin Luo1ABCDEFG, Yaqin Wang1ABCDEFG, Qianrong Qi1CE, Yan Cheng1BD, Wangming Xu1A, Jing Yang1AG*DOI: 10.12659/MSM.927709
Med Sci Monit 2021; 27:e927709






Abstract
BACKGROUND: This study aims to explore the effect of Sinomenine (SIN) on pregnancy outcomes of recurrent spontaneous abortion (RSA) in a mouse model.
MATERIAL AND METHODS: Thirty female CBA/J mice were allocated into 3 groups randomly, then mated with BALB/c mice (CBA/J×BALB/c) as normal-pregnancy group (n=10), or mated with DBA/2 mice (CBA/J×DBA/2) as RSA model (n=10), or CBA/J×DBA/2 mice treated with SIN as RSA+SIN group (n=10). The number of surviving and reabsorbed embryos in each group were counted on day 13.5 of gestation. The mouse serum was collected to determine the levels of interferon-γ (IFN)-γ and IL-4 by ELISA. Immunohistochemistry, qRT-PCR and immunoblotting were used to determine the location, mRNA and protein expressions of IFN-γ, IL-4, T-bet and GATA3 in the decidual and placental tissue.
RESULTS: In the RSA group, the amount of reabsorbed embryo was significantly higher than that in the normal-pregnancy group. However, SIN treatment showed a rescue effect on spontaneous abortion in RSA mice. IFN-γ, IL-4, T-bet, and GATA3 were all expressed in placental tissues and mainly located in the cytoplasm. The RSA group demonstrated higher expression levels of IFN-γ and T-bet than in the RSA+SIN and normal-pregnancy groups. Although RSA and RSA+SIN groups showed lower expression levels of IL-4 and GATA3 than in the normal-pregnancy group, there was no significant difference between RSA and RSA+SIN groups regarding IL-4 and GATA expression levels.
CONCLUSIONS: SIN treatment demonstrates a therapeutic effect on spontaneous abortion in RSA mice, possibly through regulating the balance of Th1/Th2 in maternal circulation and decidual tissues.
Keywords: Abortion, Habitual, Sinomenium, Th1-Th2 Balance, Abortion, Spontaneous, Decidua, Embryo, Mammalian, GATA3 Transcription Factor, interferon-gamma, Interleukin-4, Morphinans, Placenta, Pregnancy, T-Box Domain Proteins
Background
In humans, recurrent spontaneous abortion (RSA) is defined as the occurrence of 2 or more consecutive spontaneous losses of a clinical pregnancy before 20 weeks of gestation or before the embryo/fetus reaches 400 grams [1]. An estimated 5% of conceived women are affected by RSA worldwide [2]. Several mechanisms have been described for the pathogenesis of RSA, including anatomic abnormalities, couples or embryonic chromosomal abnormalities, endocrine problems, hormonal causes, and infections [3]. Notably, Th1/Th2 cytokines imbalance has been proposed to contribute to the pathogenesis of RSA [4].
In the early stage of pregnancy, fetal tolerance by maternal immune cells is vital to the maintenance of pregnancy [2]. The embryo expresses paternal antigens foreign to the mother and therefore may be viewed as an allograft, but in normal pregnancy, the embryo does not undergo maternal immune rejection. T lymphocytes, especially CD4+T helper (Th) cells, are mainly responsible for secreting cytokines that direct the immune cells to attack abnormal or infected cells. Based on their profiles of cytokine production and functions, Th cells can be classified into 2 major subsets: Th1 and Th2 cells. Both Th1 and Th2 cells have a critical role in immune allograft rejection or tolerance. The elevation of Th2-type cytokines like interleukin-4 (IL-4), localized at the feto-maternal interface, favors the maintenance of mammalian pregnancy, while Th1-type cytokines such as interferon-gamma (IFN-γ) mediate fetal rejection. It is suggested that activated Th1-type response is associated with RSA [5,6]. Several immune therapeutic methods have been used in clinical practice to prevent or treat women with unexplained RSA. However, the therapeutic effect and underlying mechanisms need to be further determined.
Many factors have been described to regulate the balance between Th1 and Th2 cytokines [7]. T-bet, a transcription factor specifically expressed in Th1 cells, is thought to induce IFN-γ production and initiate Th1 development [8]. Overexpression of T-bet gene by retroviral transfection induces IFN-γ and represses IL-4 and IL-5 production in Thp, Th1, and Th2 cells [9]. In contrast, GATA-3, a zinc finger protein, is selectively expressed in Th2 cells and is essential for the transcriptional regulation of all Th2 cytokines. Overexpression of GATA3 by retroviral transfection stimulates IL-4 and IL-5 and suppresses IFN-γ production in Th1 cells [10]. It has been shown that the upregulation of T-bet and downregulation of GATA-3 are involved in RSA [11]. T-bet is known to activate Th1 development, while GATA-3 is essential for Th2 development. Therefore, the ratio of T-bet/GATA-3 theoretically can be used as a marker to evaluate the Th1/Th2 immune balance in RSA.
Material and Methods
ANTIBODIES AND REAGENTS:
SIN was purchased from Shanghai Winherb Medical S&T Development Co., Ltd. (Shanghai, China). ELISA kits for IFN-γ and IL-4 were obtained from R&D Systems (Minneapolis, MN, USA). Anti-IFN-γ, anti- IL-4, anti-Tet and, anti-GATA-3, and GAPDH antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Horseradish peroxidase-conjugated goat anti-mouse IgG and goat anti-rabbit IgG were obtained from Bio-Rad Laboratories, Inc. (Hercules, CA, USA).
RSA MOUSE MODEL ESTABLISHMENT AND DRUG TREATMENT:
Male BALB/c mice aged 8–10 weeks were purchased from Hubei Center for Disease Control and Prevention (Wuhan, China). Female CBA/J mice and male DBA/2 mice were provided by Beijing HFK Bioscience Co., Ltd. (Beijing, China). Animals were maintained under regular laboratory conditions (12 h light: 12 h darkness cycle at 25°C to 27°C). All the processes and experiments were authorized by the Ethics Review Committee of Wuhan University Medical College.
Thirty female CBA/J mice were randomly allocated into 3 groups (n=10/group). Ten female CBA/J mice were mated with 5 male BALB/c mice to create a normal-pregnancy group (control group); 10 female CBA/J mice were mated with 5 male DBA/2 males to create an RSA group; and 10 female CBA/J mice were mated with 5 male DBA/2 males then orally administered with 100 mg/kg SIN from the 0.5 days after pregnancy to create an RSA+SIN group. The Control group and RSA group were administered the same volume of saline for 13 days. The initiation of pregnancy was marked by the presence of a postcoital vaginal plug (Day 0.5 of pregnancy).
SAMPLE COLLECTION:
Blood samples were collected from female CBA/J mice in each group via ophthalmic venous plexus on day 13.5 of pregnancy. The serum was extracted by centrifugation at 1500×g at 4°C for 30 min and stored at −80°C. After blood was collected, the female mice were sacrificed by cervical dislocation and the numbers of surviving and reabsorbed embryos were counted. The abortion embryos were manifested as a smaller size of implantation sites and the appearance of necrosis and hemorrhage. The embryo resorption rate (R)=reabsorbed embryos/(surviving embryos+reabsorbed embryos)×100%. The single placental unit, together with the deciduas, were washed in 0.9% saline to remove the blood, and then some of the samples were immediately immersed in liquid nitrogen and stored at −70°C for western blot and quantitative real-time PCR (qRT-PCR) analysis, while the rest of the tissues were fixed by 4% paraformaldehyde, dehydrated, and embedded in paraffin.
ENZYME-LINKED IMMUNOSORBENT ASSAY (ELISA):
ELISA was performed by using mouse IFN-γ and IL-4 kits (R&D Systems) following the manufacturer’s instructions.
IMMUNOHISTOCHEMISTRY:
After being deparaffinized and rehydrated, the sections of placental tissues were incubated with specific antibodies to IFN-γ, IL-4, Tet, and GATA-3 overnight at 4°C. After washing 3 times, sections were incubated with biotin-conjugated secondary antibody for 30 min, followed by 3,3-diaminobenzidine (DAB, Wuhan Google Biosynthesis Biotechnology, Wuhan, China) staining and hematoxylin counterstaining. Five random fields were observed and photographed. Image J software was used to analyze the histochemistry score (H-score) and the average value of 5 high-power fields was used as the representative value in each group.
QRT-PCR ANALYSIS:
Cellular mRNA was extracted from decidual and placental tissues using Trizol extraction reagents (Invitrogen, Carlsbad, CA, USA). RNA (5 μg) in each group was reverse-transcribed to cDNA using the RevertAid™ First-Strand cDNA Synthesis kit (Fermentas, USA). For PCR reactions, specific primers were designed by computer assistance according to the gene bank as follows:
Real-time PCR was performed using the universal PCR master mix reagent (TaKaRa, Japan) according to the manufacturer’s instructions on a qRT-PCR instrument (ABI7500; Applied Biosystems, USA). A total of 20 μL reaction contained 10 μL 2×SYBR Premix Ex TapTM, 2 μL 2.5 mM primer, 0.4 μL 50×ROX Reference Dye, 1 μL DNA templates, and 6.6 μL ddH2O. All the experiments were repeated 3 times. The 2−ΔΔCT method was used to compare the mRNA expression level between the 2 groups.
WESTERN BLOT ANALYSIS:
Decidual and placental tissues were lysed for total protein extraction and quantified by the BCA protein assay kit (Beyotime, Jiangsu, China). We separated 60 μg of total protein from each sample on SDS-PAGE gel electrophoresis, followed by transfer onto a nitrocellulose membrane (Bio-Rad; Hercules, CA, USA). After blocking, the membranes were incubated overnight with the indicated primary antibodies at 4°C. Then, the membranes were washed 3 times and incubated with horseradish peroxidase-labeled secondary antibody (Boster Biotechnology Co., Ltd., Wuhan, China). The immunoreactive bands were detected by using enhanced chemiluminescence (ECL; Beyotime, Jiangsu, China) reagent. BandScan was used for gray-value analysis and the desired protein expression was normalized with house-keeping gene expression.
STATISTICAL ANALYSIS:
Experimental data are expressed as mean±S.E.M. The differences among groups were analyzed by ANOVA or Fisher’s exact test using the SPSS 19.0 software package. A value of
Results
EFFECT OF SIN ON EMBRYO ABORTION:
The embryo resorption rates were calculated on day 13.5 of pregnancy in the control, RSA, and RSA+SIN groups. The results showed that the embryo resorption rate in the RSA model group was substantially higher compared to that in the control group (17.21% vs. 4.92%), suggesting that the RSA mouse model was successfully established. However, the RSA+SIN group showed a similar embryo resorption rate as the control group, and had a significantly lower embryo resorption rate than in the RSA group (4.65% vs. 17.21%) (Figure 1).
EFFECT OF SIN ON SERUM LEVELS OF SPECIFIC ANTIGEN IFN-γ AND IL-4:
IFN-γ and IL-4 are inflammatory cytokines of Th1 and Th2, respectively [20]. Compared with the control group, the RSA and RSA + SIN groups showed higher serum IFN-γ level and lower serum IL-4 level. The serum IFN-γ level was significantly lower in the RSA+SIN group than in the RSA group. However, no difference was observed between the 2 groups in the serum level of IL-4 (Table 1).
EFFECT OF SIN ON EXPRESSIONS OF IFN-γ AND IL-4 IN DECIDUAL AND PLACENTAL TISSUES:
To observe the effect of SIN on the expression of IFN-γ and IL-4 in the placenta, immunohistochemistry was performed to analyze the location of IFN-γ and IL-4 in placental tissue. Both IFN-γ and IL-4 were expressed in the placenta tissues of the control and RSA mice, which were mainly located in the cytoplasm (Figure 2A). The positive expression intensities of IFN-γ and IL4 in each group were analyzed using H-score. The results indicated that the H-score of IFN-γ in the RSA group was clearly higher, while the H-score of IL-4 was dramatically lower than in the control group. SIN treatment did not alter the H-score of IL-4 compared to the RSA group, but significantly reduced the IFN-γ expression and IFN-γ/IL-4 ratio (Figure 2B). The expression of IFN-γ and IL-4 mRNA and protein in decidual and placental tissues were determined by using qRT-PCR and immunoblotting (Figure 3). Compared to the control group, the mRNA and protein expression levels of IFN-γ were substantially lower, and the expression of IL-4 was considerably lower in the RSA group. However, mRNA and protein levels of IFN-γ were drastically lower in the RSA+SIN group compared to the RSA group, but there was no significant change in the expression of IL-4. We also calculated the ratio of IFN-γ/IL-4 among the 3 groups, showing that the IFN-γ/IL-4 ratio in the RSA group was notably higher than in the control and RSA+SIN groups.
EFFECT OF SIN ON EXPRESSIONS OF T-BET AND GATA-3 IN DECIDUAL AND PLACENTAL TISSUES:
Previous studies have identified that T-box expressed in T cells (T-bet) and GATA-binding protein 3 (GATA-3) are important transcription factors for Th1 and Th2 cell differentiation [8,21]. Immunohistochemistry results showed that the protein localization of T-bet and GATA-3 were mainly in the cytoplasm of placental tissue (Figure 4A). Compared with the normal group, the H-score of T-bet was significantly higher and GATA-3 was lower in the RSA group. Notably, the H-score of T-bet was remarkably lower in the RSA+SIN group than in the RSA group, but GATA-3 showed no clear difference between the 2 groups, leading to a significant decrease in the ratio of T-bet/GATA-3 in the RSA+SIN group (Figure 4B). Similarly, the mRNA and protein level of T-bet in RSA mice was higher in the RSA group than in the control group. The results show that SIN treatment can significantly reduce T-bet expression and the ratio of T-bet/GATA-3 (Figure 5).
Discussion
In this study, we mated female CBA/J mice with male DBA/2 mice to establish an RSA mouse model. Compared to the normal-pregnancy mice, RSA mice had more resorbed embryos on day 13.5 of pregnancy, possibly due to the imbalance between Th1 and Th2 cytokines. More specifically, Th1 factors of IFN-γ and T-bet were increased, while Th2 factors of IL-4 and GATA3 were decreased in RSA mice. SIN treatment can reduce embryo absorption rate, IFN-γ level, IFN-γ/IL-4 ratio, T-bet level, and T-bet/GATA3 ratio in RSA mice, suggesting that SIN might be an effective therapeutic method for RSA by correcting Th1/Th2 balance.
A successful pregnancy requires complex mechanisms to maintain the balance between immune tolerance and effector immunity. The imbalance of this regulation system will lead to poor reproductive outcomes. A deleterious role has been attributed to Th1 cells in pregnancy because some Th1-dependent effector mechanisms play a central role in acute allograft rejection. In addition, a predominantly Th2-type immunity seems to be critical for the induction of maternal tolerance toward fetal alloantigens during pregnancy. The Th1/Th2 balance in circulation has been reported to shift to Th2 dominance during normal pregnancy, whereas Th1 dominance is associated with RSA [17,18]. However, adjusting the Th1/Th2 balance can significantly improve pregnancy outcomes in RSA patients [20,22].
In the present study, the RSA model is established by using female CBA/J mice mated with male DBA/2 mice, which shares many common features with human RSA [23]. The higher spontaneous abortion rate in these RSA mice has provided a valuable model for exploring mechanisms that have been validated by studying patients with recurrent miscarriage [24,25]. Our results demonstrate that the RSA group had a higher rate of embryo resorption than in the control group. A higher concentration of Th1 cytokine (IFN-γ) corresponds with higher levels of transcription factor T-bet expression, while lower Th2 cytokine (IL-4) expression corresponds with the lower levels of GATA-3 expression observed in RSA mice. Our results showed that an RSA mouse model was successfully established, and this further confirmed that the high rates of embryo reabsorbed in the RSA mice were associated with Th1/Th2 imbalance.
SIN has a wide range of pharmacological profiles; therefore, several molecular mechanisms may be involved in its beneficial therapeutic effects. Studies have shown that SIN suppresses the activities of lymphocytes by mainly inhibiting their proliferation, inducing the apoptosis of T lymphocytes, and attenuating the Th1/Th2 imbalance [17–19]. Based on its potent anti-inflammatory properties, SIN was used for the treatment of RSA in this study. At the beginning of the study, we first used different doses (50, 100, and 150 mg/kg) of SIN to treat RSA mice for preliminary experiments and found that when the dose of SIN was 100 mg/Kg and 150 mg/kg, the embryo loss rates of RSA mice were significantly lower (data not shown). Therefore, the lowest effective dose, 100 mg/Kg, was chosen for further experiments. Our results have shown that SIN can practically inhibit T-bet expression as well as IFN-γ production in deciduas and serum of RSA mice, while it does not affect the expression levels of GATA-3 and IL-4. Since T-bet is a Th1 transcription factor and GATA-3 is a Th2 transcription factor, the results suggest that SIN can regulate Th1 immune response, but has no significant effect on the regulation of Th2 activation, which is in agreement with a previous study by Cheng et al. [18]. Taken together, the evidence suggests that SIN may be an effective immunosuppressive agent for the treatment of RSA.
However, there are still several limitations in this study. First, in addition to IFN-γ and IL-4, there are many other Th1 and Th2 cytokines, such as TNF-α, IL-5, and IL-10. Due to insufficient budget, in this study, only IFN-γ and IL-4, which are closely related to SIN, were selected as observation indicators according to previous reports. Second, the efficacy of different administration methods was not compared. In the next step of our research, we will expand the sample size, improve the detection indicators, explore the best way of administration, and use
Conclusions
This study demonstrated that SIN has a significant therapeutic effect on spontaneous abortion in RSA mice, possibly through regulating the balance of Th1/Th2 in maternal circulation and decidual tissues, which might provide a new therapeutic method for RSA treatment.
Figures
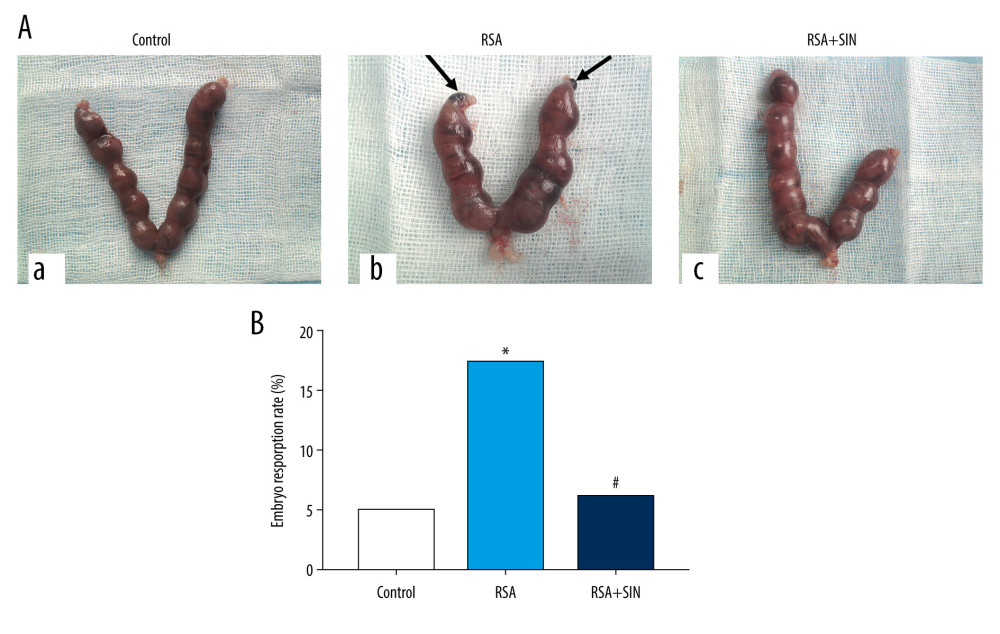 Figure 1. Effect of SIN on embryo abortion in RSA mice. (A) Embryo abortion sites of pregnant mice on Day 13.5 of pregnancy in each group. a: control group, 0 embryos resorbed; b: RSA group, 2 embryos lost; c: SIN-treated group, 0 embryo resorbed. Arrows indicate resorbed embryos. (B) Comparison of embryo resorption rates in each group. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.
Figure 1. Effect of SIN on embryo abortion in RSA mice. (A) Embryo abortion sites of pregnant mice on Day 13.5 of pregnancy in each group. a: control group, 0 embryos resorbed; b: RSA group, 2 embryos lost; c: SIN-treated group, 0 embryo resorbed. Arrows indicate resorbed embryos. (B) Comparison of embryo resorption rates in each group. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group. 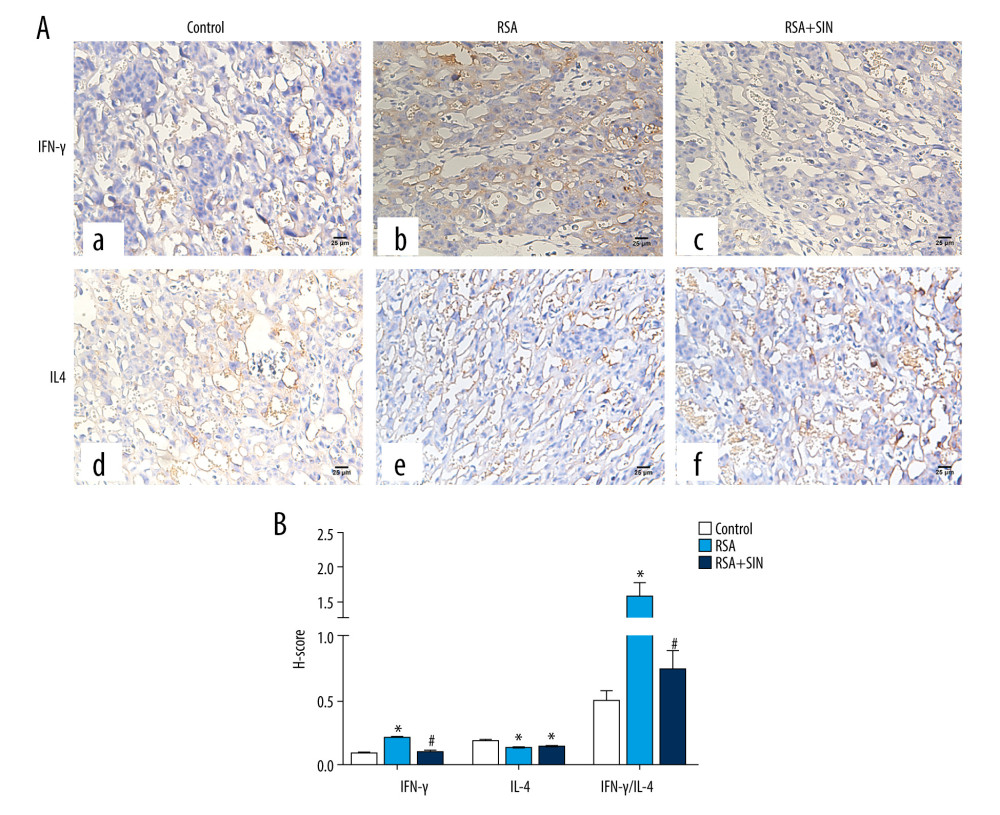 Figure 2. Effect of SIN on IFN-γ and IL-4 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of IFN-γ (a–c) and IL- 4 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of IFN-γ, IL-4, and IFN-γ/IL-4 ratio. Data are expressed as mean±S.E.M.* P<0.05, compared with the control group; # P<0.05, compared with the RSA group.
Figure 2. Effect of SIN on IFN-γ and IL-4 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of IFN-γ (a–c) and IL- 4 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of IFN-γ, IL-4, and IFN-γ/IL-4 ratio. Data are expressed as mean±S.E.M.* P<0.05, compared with the control group; # P<0.05, compared with the RSA group. 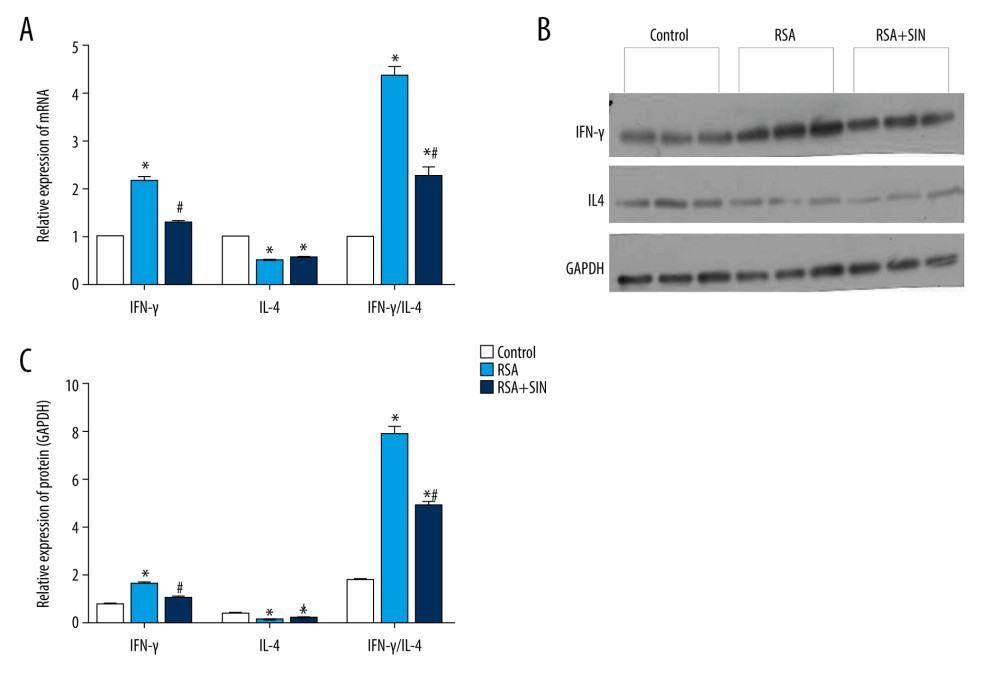 Figure 3. Effect of SIN on expressions of IFN-γ and IL-4 in decidual and placental tissues of RSA mice. (A) The mRNA levels of IFN-γ and IL-4 in the control, RSA, and SIN-treated mice were examined using RT-qPCR, and the IFN-γ/IL-4 ratio was calculated. (B) The protein expression levels of IFN-γ and IL-4 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for IFN-γ, IL-4, and IFN-γ/IL4 ratio. Data are presented as mean±SEM. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.
Figure 3. Effect of SIN on expressions of IFN-γ and IL-4 in decidual and placental tissues of RSA mice. (A) The mRNA levels of IFN-γ and IL-4 in the control, RSA, and SIN-treated mice were examined using RT-qPCR, and the IFN-γ/IL-4 ratio was calculated. (B) The protein expression levels of IFN-γ and IL-4 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for IFN-γ, IL-4, and IFN-γ/IL4 ratio. Data are presented as mean±SEM. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group. 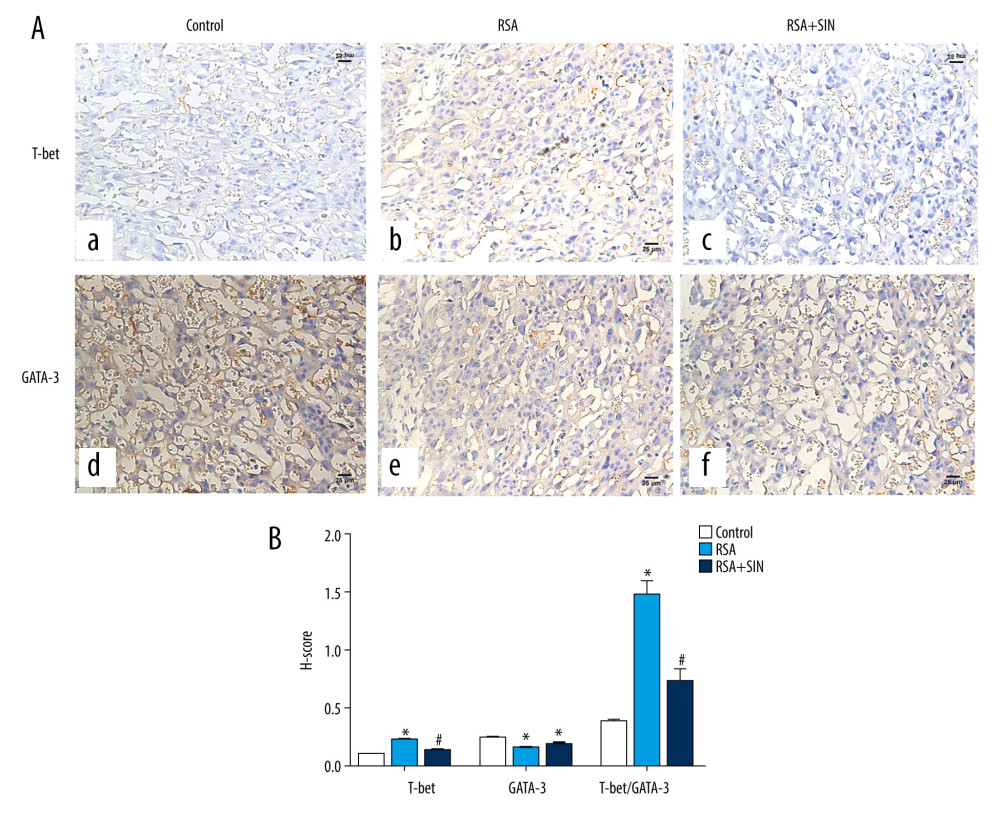 Figure 4. Effect of SIN on T-bet and GATA-3 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of T-bet (a–c) and GATA-3 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are expressed as mean±S.E.M. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.
Figure 4. Effect of SIN on T-bet and GATA-3 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of T-bet (a–c) and GATA-3 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are expressed as mean±S.E.M. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group. 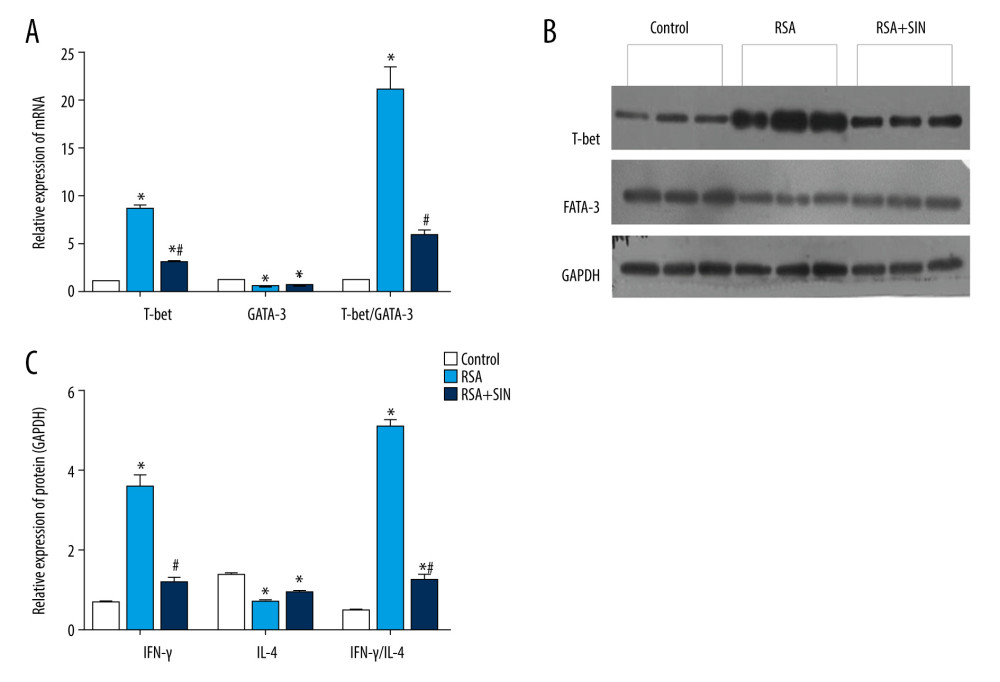 Figure 5. Effect of SIN on expressions of T-bet and GATA-3 in decidual and placental tissues of RSA mice. (A) The mRNA levels of T-bet and GATA-3 were examined using RT-qPCR and the T-bet/GATA-3 ratio was calculated. (B) The protein expression levels of T-bet and GATA-3 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are presented as mean±SEM. * P<0.05, compared with Control group; # P<0.05, compared with RSA group.
Figure 5. Effect of SIN on expressions of T-bet and GATA-3 in decidual and placental tissues of RSA mice. (A) The mRNA levels of T-bet and GATA-3 were examined using RT-qPCR and the T-bet/GATA-3 ratio was calculated. (B) The protein expression levels of T-bet and GATA-3 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are presented as mean±SEM. * P<0.05, compared with Control group; # P<0.05, compared with RSA group. 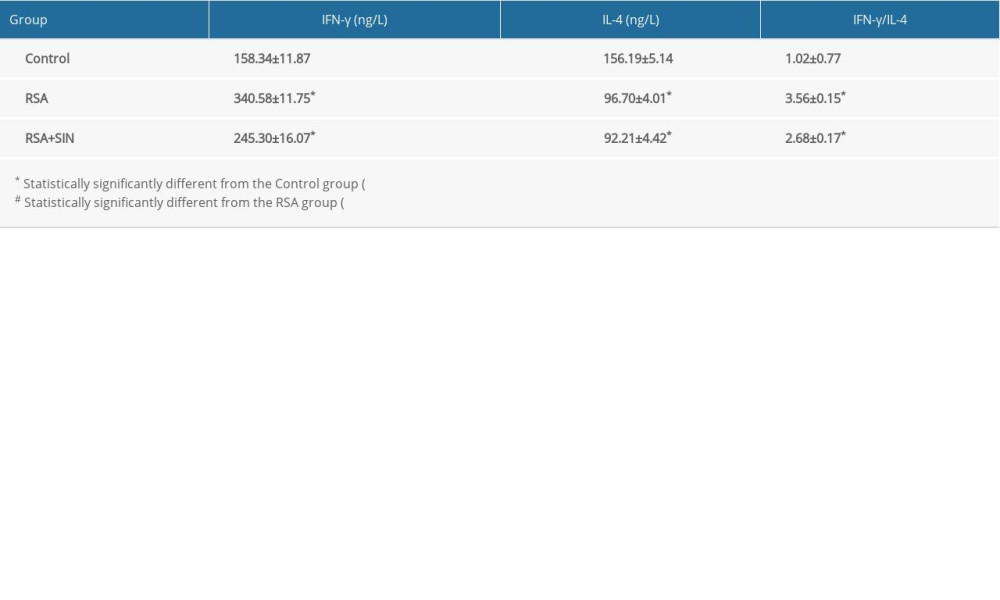
References
1. Toth B, Jeschke U, Rogenhofer N, Makrigiannakis A, Recurrent miscarriage: Current concepts in diagnosis and treatment: J Reprod Immunol, 2010; 85(1); 25-32
2. Ali S, Majid S, Niamat Ali M, Taing S, Evaluation of T cell cytokines and their role in recurrent miscarriage: Int Immunopharmacol, 2020; 82; 106347
3. Akbari S, Shahsavar F, Karami R, Recurrent spontaneous abortion (RSA) and maternal KIR genes: A comprehensive meta-analysis: JBRA Assist Reprod, 2020; 24(2); 197-213
4. Wang SW, Zhong SY, Lou LJ, The effect of intravenous immunoglobulin passive immunotherapy on unexplained recurrent spontaneous abortion: A meta-analysis: Reprod Biomed Online, 2016; 33(6); 720-36
5. Liang PY, Yin B, Cai J: Am J Reprod Immunol, 2015; 73(1); 12-21
6. Yuan J, Li J, Huang SY, Sun X, Characterization of the subsets of human NKT-like cells and the expression of Th1/Th2 cytokines in patients with unexplained recurrent spontaneous abortion: J Reprod Immunol, 2015; 110; 81-88
7. Ruterbusch M, Pruner KB, Shehata L, Pepper M: Annu Rev Immunol, 2020; 38; 705-25
8. Szabo SJ, Kim ST, Costa GL, Pillars article: A novel transcription factor, T-bet, directs Th1 lineage commitment (Cell, 2000; 100: 655–69): J Immunol, 2015; 194(7); 2961-75
9. Robinson DS, Lloyd CM, Asthma: T-bet – a master controller?: Curr Biol, 2002; 12(9); R322-24
10. Zheng WP, Flavell RA, Pillars article: The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. (Cell, 1997; 89: 587–96): J Immunol, 2016; 196(11); 4426-35
11. Qiu LH, Lin QDStudy on the expression of transcription factor GATA-3 and T-bet mRNA in decidua of women with unexplained recurrent spontaneous abortion: Zhonghua Fu Chan Ke Za Zhi, 2007; 42(2); 96-98 [in Chinese]
12. Gao LN, Zhong B, Wang Y, Mechanism underlying antitumor effects of sinomenine: Chin J Integr Med, 2019; 25(11); 873-78
13. Shen J, Yao R, Jing M, Zhou Z, Sinomenine regulates inflammatory response and oxidative stress via nuclear factor kappa B (NF-kappaB) and NF-E2-related factor 2 (Nrf2) signaling pathways in ankle fractures in children: Med Sci Monit, 2018; 24; 6649-55
14. Zhang X, Rui L, Wang M, Sinomenine attenuates chronic intermittent hypoxia-induced lung injury by inhibiting inflammation and oxidative stress: Med Sci Monit, 2018; 24; 1574-80
15. Feng ZT, Yang T, Hou XQ, Sinomenine mitigates collagen-induced arthritis mice by inhibiting angiogenesis: Biomed Pharmacother, 2019; 113; 108759
16. Wang Q, Li XK, Immunosuppressive and anti-inflammatory activities of sinomenine: Int Immunopharmacol, 2011; 11(3); 373-76
17. Gu B, Zeng Y, Yin C, Sinomenine reduces iNOS expression via inhibiting the T-bet IFN-gamma pathway in experimental autoimmune encephalomyelitis in rats: J Biomed Res, 2012; 26(6); 448-55
18. Cheng Y, Zhang J, Hou W, Immunoregulatory effects of sinomenine on the T-bet/GATA-3 ratio and Th1/Th2 cytokine balance in the treatment of mesangial proliferative nephritis: Int Immunopharmacol, 2009; 9(7–8); 894-99
19. Feng H, Yamaki K, Takano H, Suppression of Th1 and Th2 immune responses in mice by Sinomenine, an alkaloid extracted from the chinese medicinal plant Sinomenium acutum: Planta Med, 2006; 72(15); 1383-88
20. Jiao X, Wang L, Wei Z, Vitamin D deficiency during pregnancy affects the function of Th1/Th2 cells and methylation of IFN-gamma gene in offspring rats: Immunol Lett, 2019; 212; 98-105
21. Hong JH, Kim SH, Lee YC: Molecules, 2019; 24(5); 852
22. Azizi R, Ahmadi M, Danaii S, Cyclosporine A improves pregnancy outcomes in women with recurrent pregnancy loss and elevated Th1/Th2 ratio: J Cell Physiol, 2019; 234(10); 19039-47
23. Clark DA, Manuel J, Lee L, Ecology of danger-dependent cytokine-boosted spontaneous abortion in the CBA x DBA/2 mouse model. I. Synergistic effect of LPS and (TNF-alpha+IFN-gamma) on pregnancy loss: Am J Reprod Immunol, 2004; 52(6); 370-78
24. Li G, Yang L, Li D, Effects of combined treatment with PDL1 Ig and CD40L mAb on immune tolerance in the CBA/J x DBA/2 mouse model: Mol Med Rep, 2020; 21(4); 1789-98
25. Chung Y, Kim H, Im E, Th 17 cells and Nesfatin-1 are associated with spontaneous abortion in the CBA/j x DBA/2 mouse model: Dev Reprod, 2015; 19(4); 243-52
Figures
Figure 1. Effect of SIN on embryo abortion in RSA mice. (A) Embryo abortion sites of pregnant mice on Day 13.5 of pregnancy in each group. a: control group, 0 embryos resorbed; b: RSA group, 2 embryos lost; c: SIN-treated group, 0 embryo resorbed. Arrows indicate resorbed embryos. (B) Comparison of embryo resorption rates in each group. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.Figure 2. Effect of SIN on IFN-γ and IL-4 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of IFN-γ (a–c) and IL- 4 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of IFN-γ, IL-4, and IFN-γ/IL-4 ratio. Data are expressed as mean±S.E.M.* P<0.05, compared with the control group; # P<0.05, compared with the RSA group.Figure 3. Effect of SIN on expressions of IFN-γ and IL-4 in decidual and placental tissues of RSA mice. (A) The mRNA levels of IFN-γ and IL-4 in the control, RSA, and SIN-treated mice were examined using RT-qPCR, and the IFN-γ/IL-4 ratio was calculated. (B) The protein expression levels of IFN-γ and IL-4 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for IFN-γ, IL-4, and IFN-γ/IL4 ratio. Data are presented as mean±SEM. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.Figure 4. Effect of SIN on T-bet and GATA-3 location in placental tissues of RSA mice. (A) Immunohistochemical staining for the expression of T-bet (a–c) and GATA-3 (d–f) in the placenta of the control (a, d), RSA (b, e) and SIN-treated (c, f) mice. Magnification: 400×. (B) H-score for the immunohistochemical staining intensity of T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are expressed as mean±S.E.M. * P<0.05, compared with the control group; # P<0.05, compared with the RSA group.Figure 5. Effect of SIN on expressions of T-bet and GATA-3 in decidual and placental tissues of RSA mice. (A) The mRNA levels of T-bet and GATA-3 were examined using RT-qPCR and the T-bet/GATA-3 ratio was calculated. (B) The protein expression levels of T-bet and GATA-3 were detected by western blot analysis. Experiments were performed in 3 independent replicates with essentially identical results, and representative results are shown. (C) Densitometer analysis results for T-bet, GATA-3, and T-bet/GATA-3 ratio. Data are presented as mean±SEM. * P<0.05, compared with Control group; # P<0.05, compared with RSA group. In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387