07 August 2020: Lab/In Vitro Research
Reelin Promotes Cisplatin Resistance by Induction of Epithelial-Mesenchymal Transition via p38/GSK3β/Snail Signaling in Non-Small Cell Lung Cancer
Ji-Min Li1ABCE, Fang Yang1C, Juan Li2C, Wei-Qi Yuan1D, Hao Wang1BG, Yi-Qin Luo1DG*DOI: 10.12659/MSM.925298
Med Sci Monit 2020; 26:e925298






Abstract
BACKGROUND: Emerging evidence suggests the involvement of Reelin in chemoresistance in various cancers. However, its function in cisplatin (DDP) sensitivity of non-small cell lung cancer (NSCLC) needs to be investigated.
MATERIAL AND METHODS: Reelin expression in cisplatin-sensitive A549 cells and cisplatin-resistant NSCLC (A549/DDP) cells was analyzed by western blot analysis. qRT-PCR, western blotting, immunofluorescence, CCK-8 assays, Annexin V/propidium iodide apoptosis assay, and Transwell migration assays were carried out to determine the function of Reelin on DDP resistance.
RESULTS: Reelin was markedly increased in A549/DDP cells relative to A549 cells. Knockdown of Reelin enhanced DDP chemosensitivity of A549/DDP cells, whereas overexpression of Reelin enhanced DDP resistance of A549, H1299, and H460 cells. Reelin induced DDP resistance in NSCLC cells via facilitating epithelial-mesenchymal transition (EMT). Furthermore, Reelin modulated p38/GSK3β signal transduction and promoted Snail (EMT-associated transcription factor) expression. Suppression of p38/Snail reversed Reelin-induced EMT and resistance of NSCLC cells to DDP.
CONCLUSIONS: These data indicated that Reelin induces DDP resistance of NSCLC by regulation of the p38/GSK3β/Snail/EMT signaling pathway and provide evidence that Reelin suppression can be an effective strategy to suppress DDP resistance in NSCLC.
Keywords: Cisplatin, Gait Disorders, Neurologic, Cell Adhesion Molecules, Neuronal, Extracellular Matrix Proteins, glycogen synthase kinase 3 beta, Nerve Tissue Proteins, Reelin Protein, Serine Endopeptidases, Snail Family Transcription Factors
Background
Lung cancer is a predominant malignancy and common cause of cancer mortality globally. Among different forms of the condition, non-small cell lung cancer (NSCLC) accounts for about 85% of all cases [1]. Cisplatin (DDP) is a component of the chemotherapeutic regimens for NSCLC and interacts with DNA to form adducts, which lead to suppression of DNA replication and subsequent cell death [2]. However, many patients with lung cancer develop cisplatin resistance, and this resistance limits the efficacy of cisplatin [3,4]. Thus, there is a need to comprehend the underlying mechanisms of chemoresistance arises to advance the therapy of NSCLC.
Epithelial-mesenchymal transition (EMT) is a vital step in embryonic development, wound healing, tumor invasion, tissue remodeling, and metastasis [5,6]. EMT is associated with increased expressions of the mesenchymal markers vimentin and N-cadherin and decreased levels of the epithelial marker E-cadherin. Recently, increasing evidence has implicated EMT in the onset of drug resistance [7,8]. EMT has been found in some drug-resistant cancer cells [9,10], and inhibition of EMT was shown to attenuate drug resistance in many types of cancer [9,11,12]. These studies demonstrated that EMT could be related to the occurrence of chemo-resistance.
Reelin is an extracellular glycoprotein regulating neuronal migration, adhesion, and positioning [13–15]. Although the physiological role of Reelin was considered to be focused in the brain, aberrant expression of Reelin was recently reported in several tumor tissues, including lymphoma [16], multiple myeloma [17], esophageal cancer [18], prostate cancer [19], and breast cancer [20]. In addition, increased Reelin expression was proved as a poor prognostic factor in multiple myeloma [21], high Gleason score prostate cancer [19], and retinoblastoma [22]. Moreover, Reelin was shown to promote the pathogenesis of non-Hodgkin lymphoma [16], multiple myeloma [17], breast cancer [23], and mammary tumors [24]. Notably, further characterization of Reelin revealed its association with drug resistance in multiple myeloma [25]. However, the relationship between Reelin and cisplatin sensitivity in lung cancer has not been investigated.
Herein, we observed that Reelin expression was dramatically enhanced in cisplatin-resistant NSCLC cells (A549/DDP cells) relative to parental A549 cells with high sensitivity to cisplatin. We also show that Reelin facilitates cisplatin resistance through p38/GSK3β/Snail/EMT signaling mediation in NSCLC cells. Collectively, these results showed that Reelin may be a possible therapeutic target for attenuating tumorigenesis and chemoresistance in cisplatin-resistant NSCLC.
Material and Methods
CELL CULTURE:
Lung cancer A549, H1299, and H460 cells were procured from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China) and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 100 U/mL of both streptomycin and penicillin. We previously established a DDP-resistant A549/DDP cells. In brief, A549 cells were exposed to 0.5 μg/mL for 3 days and cells were allowed to restore in the next 3 days. And after 3 cycles, the concentrations of DDP were stepwise increased to be 1, 2, 4, 8, and 10 μg/L. Nine months later, the surviving cisplatin-resistant cells were designated as A549/DDP cells. These cell lines were grown under 5% CO2 at 37°C.
REAGENTS AND ANTIBODIES:
SB203580 and Licl was purchased from MedChem Express (Monmouth Junction, NJ, USA). Cisplatin was obtained from Sigma-Aldrich (St. Louis, MO, USA). Primary antibodies for p-c-Jun N-terminal kinases (p-JNK), phospho (p)-AKT, p-extracellular regulated protein kinases (p-ERK), p-p38, Snail, and β-catenin were obtained from Cell Signaling Technology (MA, USA). Antibodies against total (t)-JNK, t-p38, t-AKT, t-ERK, GAPDH, β-actin, and E-cadherin were obtained from Proteintech Group (Chicago, IL, USA). Antibody against Reelin was obtained from Abcam (MA, USA).
RNA INTERFERENCE AND PLASMID TRANSFECTION:
A negative control siRNA and siRNAs against Reelin and snail were obtained from RIBOBIO (Guangzhou, China). A549/DDP cells were plated in 6-well plates and siRNAs (50 nM) was transiently transfected using Lipofectamine 3000 (CA, USA) in line with the protocol provided by the manufacturer. To generate stable A549 cell lines stably expressing Reelin or controls, A549 cells transfected with pCrl/control vector pcDNA3.0 were grown in medium with 400 μg/mL G418. Stable cell clones were identified by evaluating Reelin expression using western blotting.
QUANTITATIVE REAL-TIME PCR:
TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) was employed to isolate total RNA from cells, and RNA was then subjected to cDNA synthesis using a PrimeScript™ Kit (TaKaRa Bio Inc., Otsu, Japan) as per the methods described by the manufacturer. The target genes and GAPDH mRNA (as the reference gene) were amplified using an ABI-7500 system (Applied Biosystems Inc., Carlsbad, CA, USA) and SYBR Green fluorescence-based assay (TaKaRa Bio Inc.). The following primers were used:
The relative levels of target genes were normalized to that of GAPDH mRNA through the 2ΔΔCq method.
CELL VIABILITY ASSAY:
Cells (5×103 cells/100 μL) were cultured on 96-well plates (Costar, Cambridge, MA, USA) at 37°C for 24 hours. Cells were subjected to increasing concentrations of DDP for 48 hours. The CCK-8 assay (Dojindo, Kumamoto, Japan) was performed to assess cell viability. Optical densities (at 450 nm) were measured for each well. All the experiments were repeated in triplicate.
CELL MIGRATION EXPERIMENTS:
The migration ability of cells was assessed using a Transwell system (8 mm; Corning Costar Co., USA) in accordance with the provider’s instructions. Cells (5×104) were resuspended in serum-free medium and placed in the top chamber of the insert; the bottom chamber was filled with DMEM supplemented with 10% FBS. After incubation for 4 hours, the cells on the lower side of the membrane were fixed, stained, and counted under a microscope.
FLOW CYTOMETRY:
Cell apoptosis was detected by staining with Annexin V/propidium iodide (KeyGEN, Nanjing, China) using flow cytometry (BD FACSCalibur) as per the methods described by the manufacturer.
IMMUNOFLUORESCENCE:
Cells on plates were fixed with ice-cold 4% paraformaldehyde, permeabilized using 0.1% TritonX-100, blocked with goat serum, and then incubated with specific antibodies (E-cadherin, N-cadherin) and Alexa Fluro 488 or Alexa Fluor 594-conjugated secondary antibodies (1: 1000 dilution). The image was taken using Confocal Laser Scanning Microscopy (Zeiss).
WESTERN BLOT ASSAY:
Cells were washed 3 times using ice-cold phosphate-buffered saline (PBS) and lysed using lysis buffer. The cell lysates were centrifuged (12 000 g) at 4°C for 20 minutes and then denatured for 10 minutes at 100°C. Protein concentration was evaluated by bicinchoninic acid (BCA) protein assay kit (Beyotime, China). The protein samples were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene difluoride (PVDF) membranes (0.45 μm). Membranes were blocked within 5% nonfat milk for 2 hours at room temperature. Membranes were then incubated with specific antibodies (1: 1000 dilution) overnight at 4°C, followed by incubation with the secondary antibodies (1: 10 000 dilution). Finally, a chemiluminescence reagent (Biosystem, China) was used to analyze the proteins.
STATISTICAL ANALYSES:
Data values are shown as mean±standard deviation (SD). Parametric data involving 2 treatments were analyzed with the unpaired Student’s
Results
PARENTAL A549 CELLS AND CISPLATIN-RESISTANT A549/DDP CELLS DIFFER IN MORPHOLOGY, PHYSIOLOGY, AND REELIN EXPRESSION:
To investigate whether Reelin regulates the cisplatin resistance of lung cancer cells, we established cisplatin-resistant A549 cells (A549/DDP cells) as described in Methods. The A549/DDP cells exhibited stronger resistance to cisplatin than that in A549 cells (Figure 1A). A549/DDP cells were spindle-shaped and detached from each other, while parental A549 cells showed a rounded shape (Figure 1B). These differences in morphology indicated that A549/DDP cells acquired a mesenchymal phenotype, which was confirmed by decreased E-cadherin expression and increased N-cadherin expression (Figures 1C–1E). Migration assays revealed that A549/DDP cells migrated faster than A549 cells (Figure 1F).
We next determined the expression of Reelin in A549 and A549/DDP cells. The results showed that the level of Reelin protein expression was considerably enhanced in A549/DDP cells relative to A549 cells (Figure 1G). To investigate the effect of chemotherapeutics on the expression of Reelin, we treated A549 cells with cisplatin (1 μg/mL) for 12, 24, and 48 hours and examined Reelin levels by western blot. As shown in Figure 1H, Reelin expression clearly increased in A549 cells after treatment with cisplatin for 12 and 24 hours. These findings indicated that Reelin might be participated in mediating chemoresistance in NSCLC cells.
REELIN ENHANCES CISPLATIN RESISTANCE IN NSCLC CELLS:
To examine the function of Reelin in cisplatin resistance, we next performed transient knockdown experiments in NSCLC cells. A549/DDP cells transfected with Reelin or control siRNA were exposed to various concentrations of cisplatin and cell viability was evaluated using Cell Counting Kit-8 (CCK-8) assay. Western blot confirmed that Reelin siRNA efficiently reduced Reelin protein expression (Figure 2A). We found that Reelin suppression reduced A549/DDP cell viability relative to control siRNA-transfected cells (Figure 2B). Furthermore, knockdown of Reelin in A549/DDP cells increased cisplatin-triggered cell apoptosis relative to control cells (Figure 2C). We also assessed the impact of Reelin upregulation on the sensitivity of A549 cells to cisplatin (Figure 2D). Reelin overexpression in A549 cells markedly enhanced their cisplatin resistance (Figure 2E) and attenuated cisplatin-induced apoptosis (Figure 2F). We further investigated the association between Reelin and cisplatin resistance in other NSCLC cells. We found that overexpression of Reelin in H1299 and H460 cells increased their resistance to cisplatin (Figure 2G). Collectively, these findings suggested that upregulation of Reelin facilitates the resistance of NSCLC cells to cisplatin.
REELIN PROMOTES EMT IN NSCLC CELLS:
We next examined the function of EMT in Reelin-mediated resistance of NSCLC cells to cisplatin. Reelin was knocked down in A549/DDP cells (A549/DDP si-Reelin) and overexpressed in A549 cells (A549/Reelin). As shown in Figure 3A, we observed EMT-induced morphological alterations in A549/Reelin cells, including irregular and elongated mesenchymal phenotype, while A549/DDP si-Reelin cells showed an epithelial phenotype. These morphological changes were verified by examination of EMT-related molecular markers. A549/Reelin cells showed overexpression of N-cadherin and reduced expression of E-cadherin, while A549/DDP si-Reelin cells exhibited the opposite trends in EMT-associated marker expression (Figure 3B–3D). Consistent with these results, Transwell assays revealed that upregulation of Reelin markedly promoted the migration of A549 cells (Figure 3E), whereas Reelin knockdown significantly suppressed A549/DDP cell migration (Figure 3F). Together these findings revealed that Reelin promotes EMT and cell migration in NSCLC cells.
REELIN ENHANCES CISPLATIN RESISTANCE IN NSCLC CELLS VIA TRIGGERING EMT:
Given that several transcription factors (i.e., Snail, Zeb1, Slug, and Twist) play crucial functions in EMT regulation [5], we examined the expression of these transcription factors in A549/Reelin cells relative to A549 cells. We observed elevated expression of Snail in A549/Reelin cells, whereas Slug, ZEB1, and Twist expressions were unchanged (Figure 4A). Furthermore, Reelin silencing resulted in significantly reduced Snail expression in A549/DDP cells (Figure 4B). However, we found that the mRNA levels of Snail showed no obvious changes in Reelin-overexpressing A549 cells or Reelin-knockdown A549/DDP cells (Figure 4C), which indicated that Reelin might regulate Snail expression at post-transcription levels. To explore the precise function of Snail in Reelin-enhanced NSCLC resistance, we transfected A549/Reelin cells with Snail siRNA and examined EMT markers and cisplatin sensitivity. Snail silencing in A549/Reelin cells induced mesenchymal-epithelial transition, occasioned by significant upregulation of E-cadherin, as well as downregulation of N-cadherin (Figure 4D), as well as decreased cell migration (Figure 4E). Notably, Snail silencing led to a considerable increase in the sensitivity of A549/Reelin cells to cisplatin (Figure 4F). Thus, these findings indicate that Reelin may promote cisplatin resistance by interacting with Snail-associated EMT in NSCLC.
P38/GSK3β PATHWAY ACTIVATION WAS INVOLVED IN REELIN-TRIGGERED EMT AND CHEMORESISTANCE IN NSCLC CELLS:
We further explored the mechanisms behind the Reelin-induced upregulation of chemoresistance, Snail expression and EMT in NSCLC cells. Previous studies showed that AKT and MAPK activation can inhibit GSK-3β [26,27], resulting in stabilization and nuclear localization of Snail, and consequently stimulate EMT as well as cell migration. Thus, we examined the expression of proteins in the AKT/MAPK/GSK-3β/Snail pathway. The results showed that Reelin overexpression significantly increased the phosphorylation of p38 and GSK3β, but reduced phosphorylation of AKT in A549 cells (Figure 5A). However, Reelin upregulation had no impact on ERK and JNK phosphorylation or β-catenin levels. In contrast, phosphorylation of p38 and GSK3β was markedly reduced in Reelin-knockdown A549/DDP cells (Figure 5B). To verify whether the tumor promoting effects of Reelin were mediated by activation of p38/GSK-3β signaling, the p38 blocker SB203580 was used to examine the association between p38/GSK-3β signaling and Reelin-induced EMT as well as cisplatin resistance. SB203580 effectively decreased Snail and p-GSK3β expressions in A549 cells overexpressing Reelin (Figure 5C). SB203580 also attenuated E-cadherin and N-cadherin expressions triggered by Reelin (Figure 5C). when we treated A549 cells with GSK-3β inhibitor LiCl, in keeping with Reelin overexpression, the expressions of Snail and p-GSK-3β were upregulated (Figure 5D). In addition, SB203580 markedly reduced cell migration and increased cisplatin cytotoxicity in Reelin-overexpressing A549 cells (Figure 5E, 5F). Together these findings suggest that Reelin induces EMT and cisplatin resistance in NSCLC through p38/GSK-3β/Snail signaling pathway activation.
Discussion
Even though current chemotherapy regimens have improved the disease outcomes of cancer patients, platinum-based chemoresistance remains a main barrier to the successful treatment of NSCLC [28]. Therefore, clarification of the potential mechanisms for chemoresistance is necessary to improve lung cancer treatment. Herein, we observed that Reelin upregulation induced chemoresistance in the A549 cells, whereas Reelin suppression selectively enhanced cisplatin cytotoxicity in the A549/DDP cells, suggesting that Reelin plays a vital function in chemoresistance related to NSCLC.
Abnormal expression of Reelin promotes tumorigenesis, including the enhancement of cell proliferation, adhesion, and invasion [16,17,25]. Previous studies have suggested a correlation between Reelin and tumor chemoresistance [25]. Overexpression of Reelin in multiple myeloma cells increased their resistance to cisplatin. Similarly, our results revealed that Reelin knockdown enhanced cisplatin cytotoxicity in A549/DDP cells, whereas Reelin overexpression promoted cisplatin resistance in A549 cells, thus implicating Reelin in NSCLC-associated cisplatin resistance.
Studies have revealed numerous mechanisms underlying chemoresistance, such as the reduction in cellular accumulation, intracellular inactivation, blocking of apoptosis induction, and enhancement of DNA repair [29,30]. Recent findings indicate that EMT may have a key role in mediating chemotherapy resistance in tumor cells [6,8]. In addition, overexpression of transcription factors of EMT, such as Twist, Slug, Snail, and ZEB, in drug-resistant cells has been related to enhanced metastatic ability and resistance to apoptosis, whereas downregulation of these transcription factors rendered cells susceptible to chemotherapy-induced apoptosis [7,10,31]. Our results showed that modulation of Reelin expression in cisplatin-sensitive and cisplatin-resistant NSCLC cells led to variations in the levels of Snail and EMT-associated proteins. Furthermore, Snail silencing successfully attenuated Reelin-induced cisplatin resistance, suggesting that the Snail-orchestrated EMT process may be a vital factor in Reelin-triggered chemoresistance of NSCLC cells. In line with our results, Reelin has been shown to promote mobility in non-Hodgkin lymphoma cells [16]. In addition, Reelin was also found to suppress the migration of hepatocellular carcinoma cells and mouse embryonic fibroblasts cells [32,33]. These results suggested the function of Reelin in tumor progression is cell type-dependent. Thus, further investigations of Reelin functions in cancer cells are warranted.
Activation of the MAPK pathway is related to tumorigenesis and has a vital function in cancer pathogenesis as it regulates cell migration, proteinase induction, and apoptosis [34–36]. We found that p38, but not ERK and JNK, showed increased phosphorylation in A549 cells in response to Reelin overexpression, while AKT phosphorylation was suppressed. These results differed from previous findings that Reelin overexpression in tumor cells promoted multiple myeloma cell adhesion and survival via AKT pathways [17], suggesting that Reelin-induced signaling may be cell type-dependent. Several studies have shown that activated p38/GSK3β signaling induced EMT and chemoresistance in cancer cells [26,28,37]. Our data showed that knockdown of Reelin result in the downregulation of p-p38 and p-GSK3β in A549/DDP cells, while upregulation of Reelin led to a dramatic increase in p-p38 and p-GSK3β levels. Notably, treatment with a p38 MAPK inhibitor attenuated the Reelin-induced upregulation of Snail in A549/Reelin cells. Therefore, modulation of Snail by Reelin may arise through p38 signaling pathway activation.
Conclusions
Together, our results show that Reelin is overexpressed in A549 cells exhibiting cisplatin-resistance. Reelin also promoted the resistance of NSCLC cells to cisplatin by stimulating p38/GSK3β/Snail/EMT signal transduction. These findings indicate that Reelin is a potential molecular target to suppress cisplatin-resistance in NSCLC.
Figures
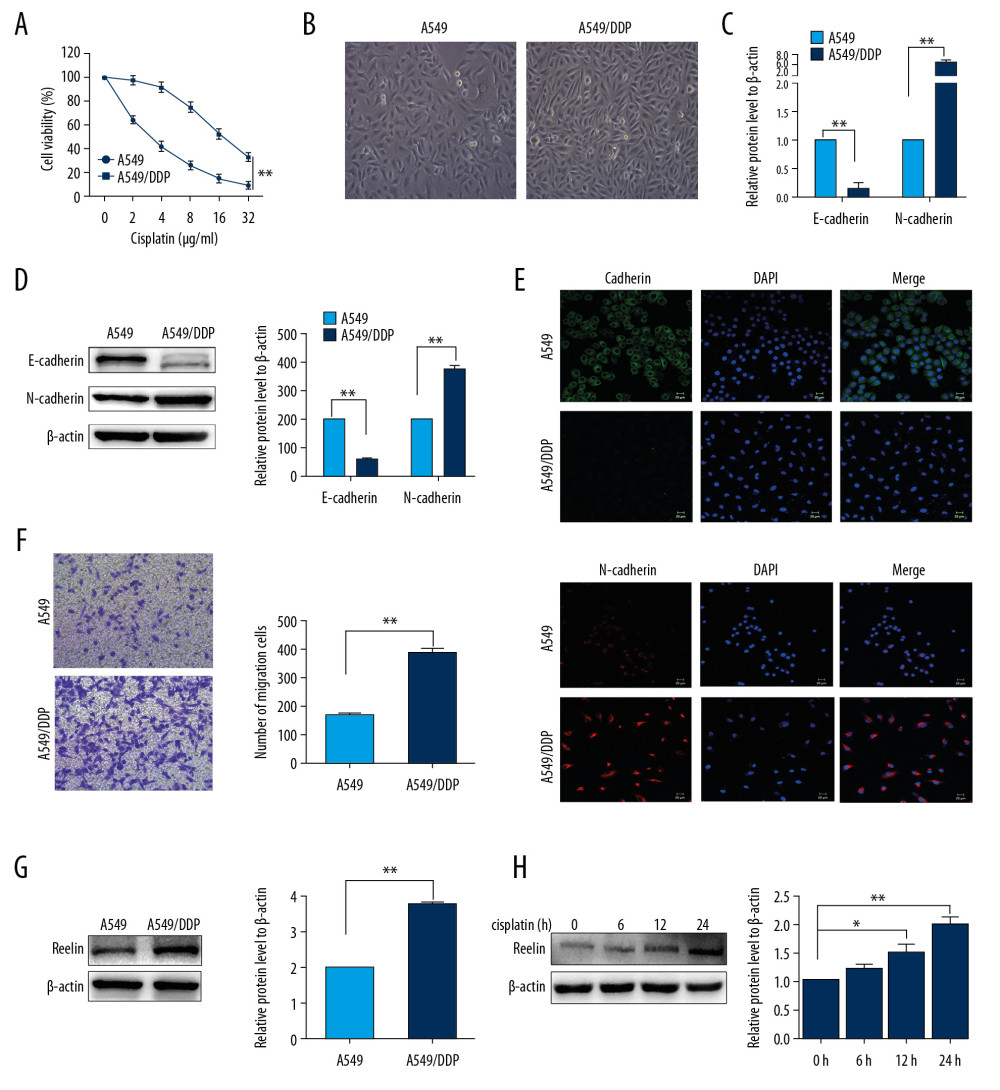 Figure 1. Differences between A549/DDP cells and parental A549 cells. (A) A549/DDP cells show increased resistance to cisplatin compared with A549 cells, as determined by CCK-8 assays. (B) A549/DDP cells exhibit a fibroblastic morphology, while A549 cells have an epithelial-like appearance. (C–E) Real-time PCR, western blot, and immunofluorescence analyses of gene and protein expression of EMT-related markers E-cadherin and N-cadherin. (F) Migration assays revealed significant enhancement of migration ability in A549/DDP cells relative to A549 cells. (G) Protein expression of Reelin was measured by western blotting. (H) A549 cells were treated with cisplatin (1 μg/mL) for the times indicated, and the protein levels of Reelin were determined by western blotting. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8; PCR – polymerase chain reaction; EMT – epithelial-mesenchymal transition.
Figure 1. Differences between A549/DDP cells and parental A549 cells. (A) A549/DDP cells show increased resistance to cisplatin compared with A549 cells, as determined by CCK-8 assays. (B) A549/DDP cells exhibit a fibroblastic morphology, while A549 cells have an epithelial-like appearance. (C–E) Real-time PCR, western blot, and immunofluorescence analyses of gene and protein expression of EMT-related markers E-cadherin and N-cadherin. (F) Migration assays revealed significant enhancement of migration ability in A549/DDP cells relative to A549 cells. (G) Protein expression of Reelin was measured by western blotting. (H) A549 cells were treated with cisplatin (1 μg/mL) for the times indicated, and the protein levels of Reelin were determined by western blotting. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8; PCR – polymerase chain reaction; EMT – epithelial-mesenchymal transition. 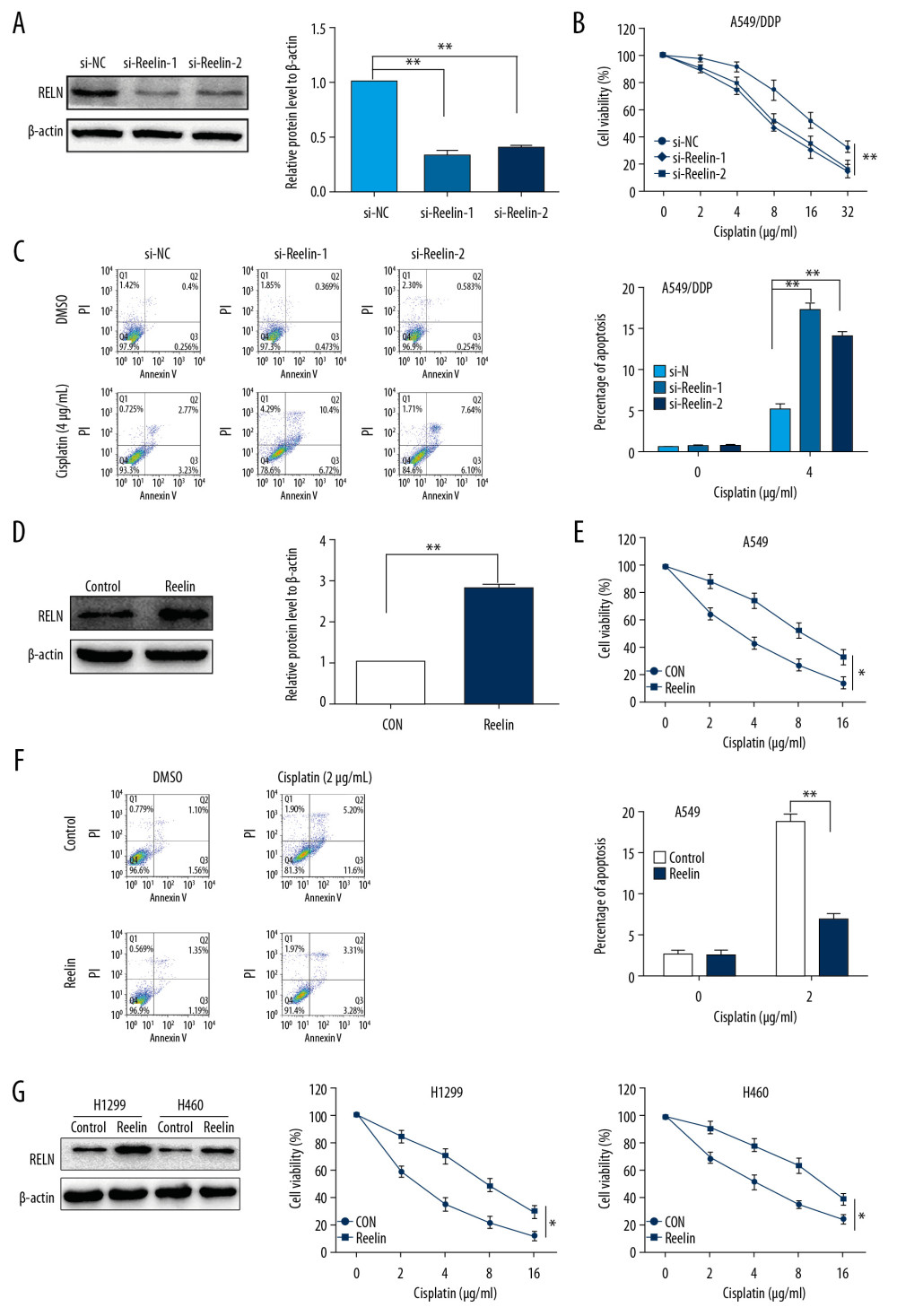 Figure 2. Reelin enhances cisplatin resistance in lung cancer cells. (A) A549/DDP cells were transfected with Reelin siRNA and Reelin expression was examined by western blotting. (B) A549/DDP cells transfected with Reelin siRNA were treated with cisplatin at the different concentrations for 48 hours; the viability was measured by CCK-8 assays. (C) A549/DDP cells were transfected with Reelin siRNA, treated with 4 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (D) A549 cells were transfected with the Reelin expression vector, and Reelin expression was evaluated by western blotting. (E) A549 cells transfected with the Reelin expression vector were treated with cisplatin at the indicated concentrations for 48 h; cell viability was determined by CCK-8 assays. (F) A549 cells were transfected with the Reelin expression vector, treated with 2 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (G) H1299 and H460 cells were transfected with Reelin, and cell viability was assessed by CCK-8 assay. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8.
Figure 2. Reelin enhances cisplatin resistance in lung cancer cells. (A) A549/DDP cells were transfected with Reelin siRNA and Reelin expression was examined by western blotting. (B) A549/DDP cells transfected with Reelin siRNA were treated with cisplatin at the different concentrations for 48 hours; the viability was measured by CCK-8 assays. (C) A549/DDP cells were transfected with Reelin siRNA, treated with 4 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (D) A549 cells were transfected with the Reelin expression vector, and Reelin expression was evaluated by western blotting. (E) A549 cells transfected with the Reelin expression vector were treated with cisplatin at the indicated concentrations for 48 h; cell viability was determined by CCK-8 assays. (F) A549 cells were transfected with the Reelin expression vector, treated with 2 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (G) H1299 and H460 cells were transfected with Reelin, and cell viability was assessed by CCK-8 assay. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8. 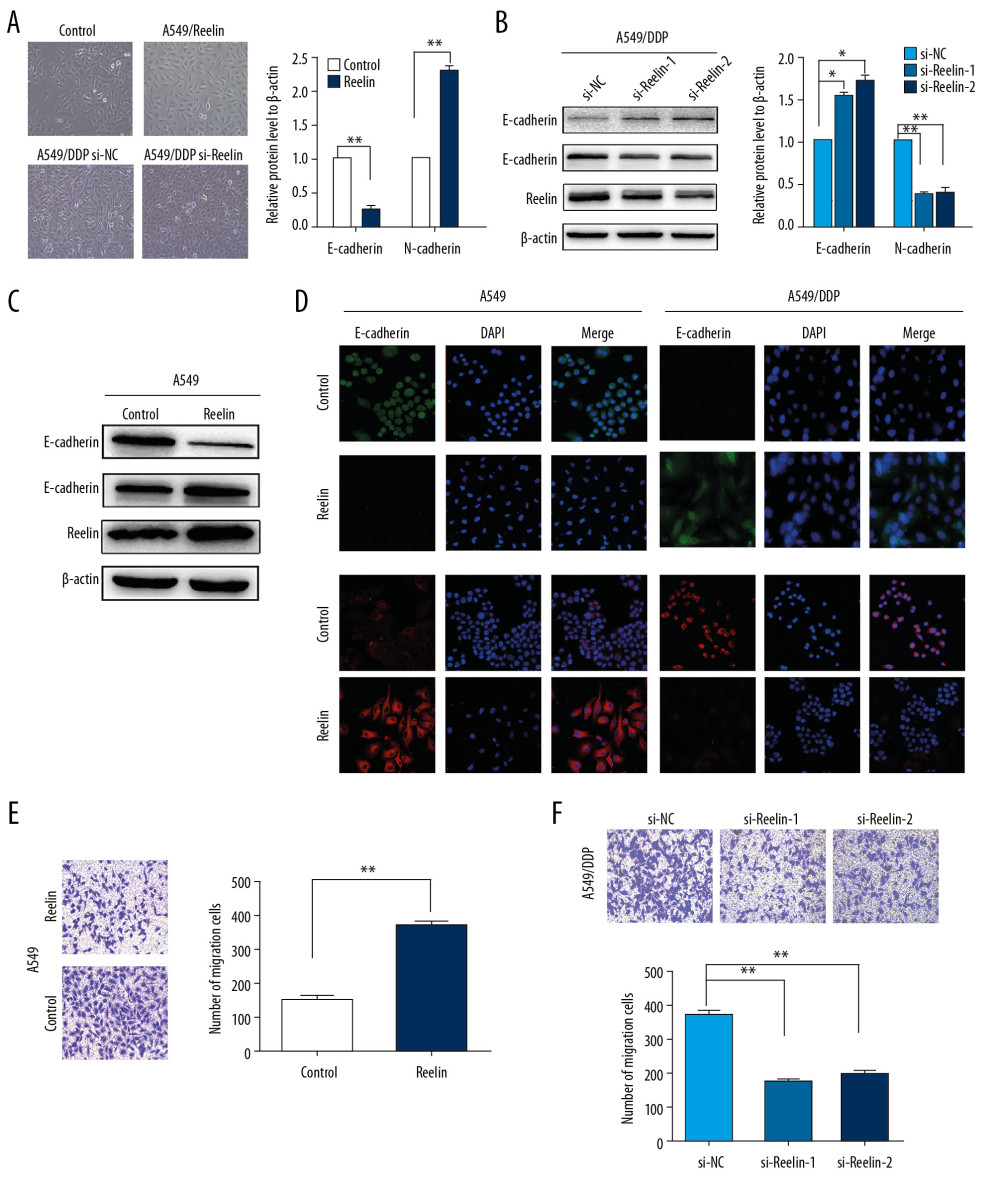 Figure 3. Reelin promotes EMT in NSCLC cells. (A) Cell morphological observations after Reelin overexpression (A549-NC versus A549/Reelin) in A549 cells and Reelin knockdown in A549/DDP cells (A549/DDP si-NC versus A549/DDP si-Reelin). (B–D) E-cadherin and N-cadherin expressions were evaluated by western blotting and immunofluorescence in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (E, F) Cell migration was measured by Transwell migration assays. * P<0.05, ** P<0.01.
Figure 3. Reelin promotes EMT in NSCLC cells. (A) Cell morphological observations after Reelin overexpression (A549-NC versus A549/Reelin) in A549 cells and Reelin knockdown in A549/DDP cells (A549/DDP si-NC versus A549/DDP si-Reelin). (B–D) E-cadherin and N-cadherin expressions were evaluated by western blotting and immunofluorescence in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (E, F) Cell migration was measured by Transwell migration assays. * P<0.05, ** P<0.01. 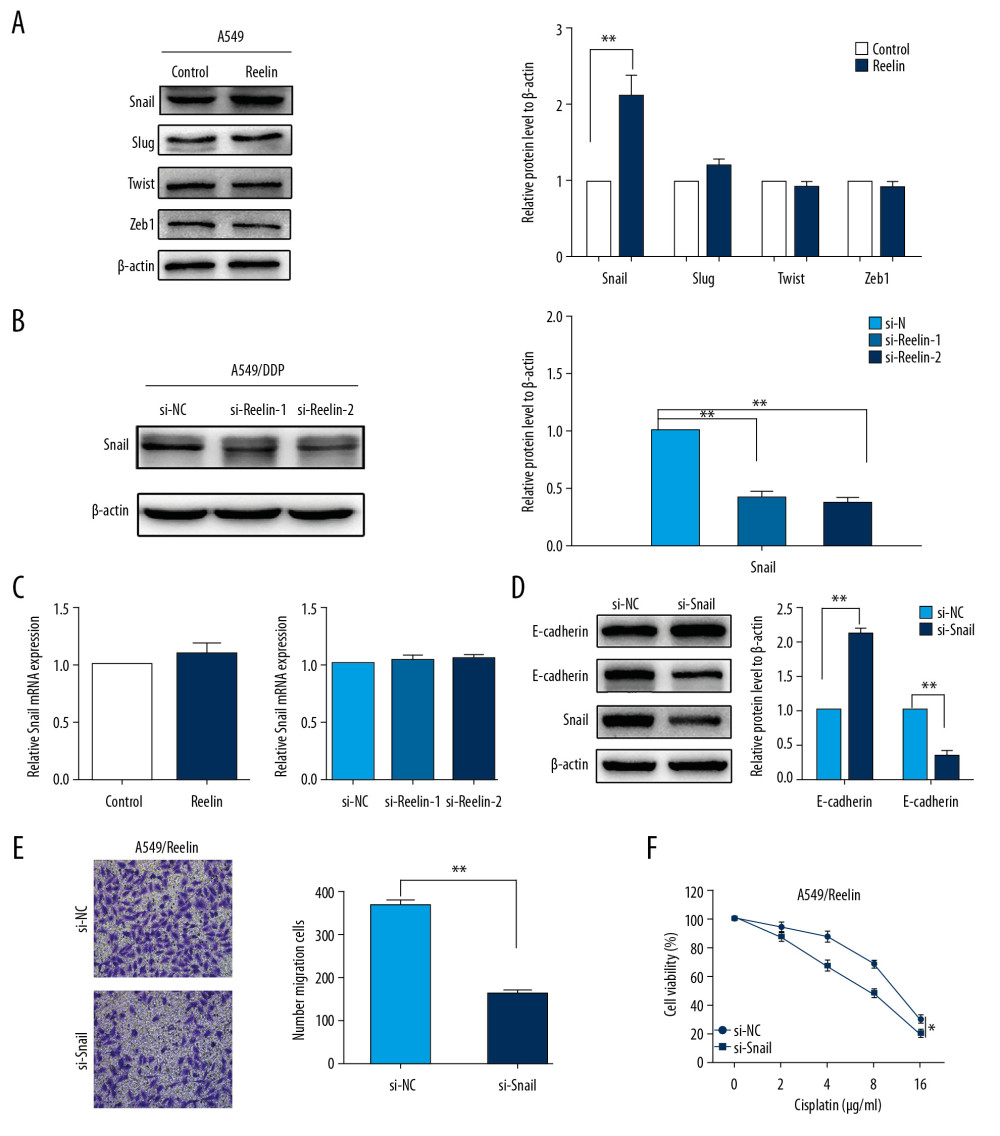 Figure 4. Silencing of Snail reverses the EMT phenotype and cisplatin resistance in NSCLC cells. (A) Effects of Reelin on Snail, ZEB1, Slug, and Twist expression in A549 cells were examined by western blotting. (B) Effects of Reelin on Snail expression in A549/DDP cells were examined by western blotting. (C) Snail expression were evaluated by qRT-PCR in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (D) The expressions of E-cadherin, N-cadherin, and Snail were evaluated by western blotting after Snail knockdown in A549/Reelin cells. (E) Transwell assays revealed dramatically increased migration ability of A549/Reelin cells transfected with si-Snail relative to those transfected with si-NC. (F) A549/Reelin cells were transfected with Snail siRNA, treated with cisplatin at the indicated concentrations for 48 hours, and measured for cell viability by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8.
Figure 4. Silencing of Snail reverses the EMT phenotype and cisplatin resistance in NSCLC cells. (A) Effects of Reelin on Snail, ZEB1, Slug, and Twist expression in A549 cells were examined by western blotting. (B) Effects of Reelin on Snail expression in A549/DDP cells were examined by western blotting. (C) Snail expression were evaluated by qRT-PCR in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (D) The expressions of E-cadherin, N-cadherin, and Snail were evaluated by western blotting after Snail knockdown in A549/Reelin cells. (E) Transwell assays revealed dramatically increased migration ability of A549/Reelin cells transfected with si-Snail relative to those transfected with si-NC. (F) A549/Reelin cells were transfected with Snail siRNA, treated with cisplatin at the indicated concentrations for 48 hours, and measured for cell viability by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8. 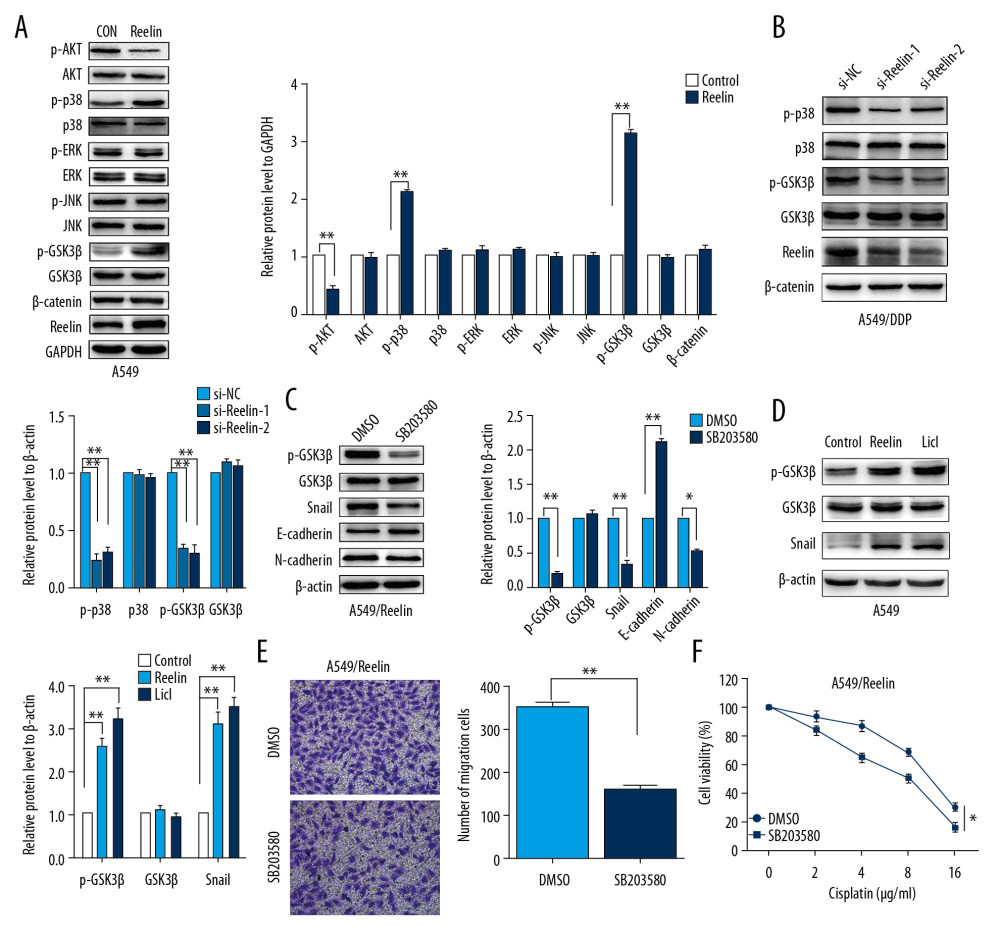 Figure 5. Activation of the p38/GSK3β signaling pathway is involved in Reelin-induced EMT and cisplatin resistance in A549 cells. (A) A549 cells were transfected with the Reelin expression vector, and p-AKT, p-p38, p-JNK, p-ERK, β-catenin, and p-GSK-3β were evaluated by western blotting. (B) A549/DDP cells were transfected with Reelin siRNA, and p-p38 and p-GSK-3β were evaluated by western blotting. (C) A549 cells transfected with the Reelin expression vector were treated with DMSO or p38 inhibitor SB203580 (10 μM) for 48 hours, and p-GSK3β, Snail, and EMT markers were evaluated by western blotting. (D) A549 cells were treated with LiCl (40 mM) for 6 hours. The expression of Snail and p-GSK-3β were analyzed by western blotting. (E) Cell migration was measured by Transwell migration assays. (F) Cell viability was analyzed by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8.
Figure 5. Activation of the p38/GSK3β signaling pathway is involved in Reelin-induced EMT and cisplatin resistance in A549 cells. (A) A549 cells were transfected with the Reelin expression vector, and p-AKT, p-p38, p-JNK, p-ERK, β-catenin, and p-GSK-3β were evaluated by western blotting. (B) A549/DDP cells were transfected with Reelin siRNA, and p-p38 and p-GSK-3β were evaluated by western blotting. (C) A549 cells transfected with the Reelin expression vector were treated with DMSO or p38 inhibitor SB203580 (10 μM) for 48 hours, and p-GSK3β, Snail, and EMT markers were evaluated by western blotting. (D) A549 cells were treated with LiCl (40 mM) for 6 hours. The expression of Snail and p-GSK-3β were analyzed by western blotting. (E) Cell migration was measured by Transwell migration assays. (F) Cell viability was analyzed by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8. References
1. Siegel RL, Miller KD, Jemal A, Cancer statistics, 2016: Cancer J Clin, 2016; 66(1); 7-30
2. Arriagada R, Bergman B, Dunant A, Cisplatin-based adjuvant chemotherapy in patients with completely resected non-small-cell lung cancer: N Engl J Med, 2004; 350(4); 351-60
3. Kim ES, Chemotherapy resistance in lung cancer: Adv Exp Med Biol, 2016; 893; 189-209
4. Galluzzi L, Vitale I, Michels J, Systems biology of cisplatin resistance: Past, present and future: Cell Death Dis, 2014; 5; e1257
5. Brabletz T, Kalluri R, Nieto MA, Weinberg RA, EMT in cancer: Nat Rev Cancer, 2018; 18(2); 128-34
6. Skarkova V, Kralova V, Krbal L, Oxaliplatin and irinotecan induce heterogenous changes in the EMT markers of metastasizing colorectal carcinoma cells: Exp Cell Res, 2018; 369(2); 295-303
7. Guo C, Ma J, Deng G, ZEB1 Promotes oxaliplatin resistance through the induction of epithelial-mesenchymal transition in colon cancer cells: J Cancer, 2017; 8(17); 3555-66
8. Singh A, Settleman J, EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer: Oncogene, 2010; 29(34); 4741-51
9. Yang AD, Fan F, Camp ER, Chronic oxaliplatin resistance induces epithelial-to-mesenchymal transition in colorectal cancer cell lines: Clin Cancer Res, 2006; 12(14 Pt 1); 4147-53
10. Wang H, Zhang G, Zhang H, Acquisition of epithelial-mesenchymal transition phenotype and cancer stem cell-like properties in cisplatin-resistant lung cancer cells through AKT/beta-catenin/Snail signaling pathway: Eur J Pharmacol, 2014; 723; 156-66
11. Ye Q, Su L, Chen D, Astragaloside IV induced miR-134 expression reduces EMT and increases chemotherapeutic sensitivity by suppressing CREB1 signaling in colorectal cancer cell line SW-480: Cell Physiol Bioch, 2017; 43(4); 1617-26
12. Ma JL, Zeng S, Zhang Y, Epithelial-mesenchymal transition plays a critical role in drug resistance of hepatocellular carcinoma cells to oxaliplatin: Tumour Biol, 2016; 37(5); 6177-84
13. Frotscher M, Cajal-Retzius cells, Reelin, and the formation of layers: Curr Opin Neurobiol, 1998; 8(5); 570-75
14. Groen JL, Ritz K, Jalalzadeh H, RELN rare variants in myoclonus-dystonia: Mov Disord, 2015; 30(3); 415-19
15. Sommen M, Van Camp G, Liktor B, Genetic association analysis in a clinically and histologically confirmed otosclerosis population confirms association with the TGFB1 gene but suggests an association of the RELN gene with a clinically indistinguishable otosclerosis-like phenotype: Otol Neurotol, 2014; 35(6); 1058-64
16. Dou A, Wang Z, Zhang N, Liu J, Loss of Reelin suppresses cell survival and mobility in non-Hodgkin lymphoma: Oncol Rep, 2017; 37(6); 3572-80
17. Qin X, Lin L, Cao L, Extracellular matrix protein Reelin promotes myeloma progression by facilitating tumor cell proliferation and glycolysis: Sci Rep, 2017; 7; 45305
18. Yuan Y, Chen H, Ma G, Reelin is involved in transforming growth factor-beta1-induced cell migration in esophageal carcinoma cells: PLoS One, 2012; 7(2); e31802
19. Perrone G, Vincenzi B, Zagami M, Reelin expression in human prostate cancer: A marker of tumor aggressiveness based on correlation with grade: Modern Pathol, 2007; 20(3); 344-51
20. Stein T, Cosimo E, Yu X, Loss of reelin expression in breast cancer is epigenetically controlled and associated with poor prognosis: Am J Pathol, 2010; 177(5); 2323-33
21. An QM, Liu YG, Lin LExpression of extracellular matrix protein reelin in patients with multiple myeloma and its relationship with prognosis: Zhongguo Shi Yan Xue Ye Xue Za Zhi, 2018; 26(3); 789-95 [in Chinese]
22. Seigel GM, Hackam AS, Ganguly A, Human embryonic and neuronal stem cell markers in retinoblastoma: Mol Vis, 2007; 13; 823-32
23. Jandial R, Choy C, Levy DM, Astrocyte-induced Reelin expression drives proliferation of Her2(+) breast cancer metastases: Clin Exp Metastasis, 2017; 34(2); 185-96
24. Khialeeva E, Chou JW, Allen DE, Reelin deficiency delays mammary tumor growth and metastatic progression: J Mammary Gland Biol Neoplasia, 2017; 22(1); 59-69
25. Lin L, Yan F, Zhao D, Reelin promotes the adhesion and drug resistance of multiple myeloma cells via integrin beta1 signaling and STAT3: Oncotarget, 2016; 7(9); 9844-58
26. Gu Y, Wang Q, Guo K, TUSC3 promotes colorectal cancer progression and epithelial-mesenchymal transition (EMT) through WNT/beta-catenin and MAPK signalling: J Pathol, 2016; 239(1); 60-71
27. Han Y, Zhang L, Wang W, Livin promotes the progression and metastasis of breast cancer through the regulation of epithelial mesenchymal transition via the p38/GSK3beta pathway: Oncol Rep, 2017; 38(6); 3574-82
28. Seve P, Dumontet C, Chemoresistance in non-small cell lung cancer: Curr Med Chem Anticancer Agents, 2005; 5(1); 73-88
29. Ferreira JA, Peixoto A, Neves M, Mechanisms of cisplatin resistance and targeting of cancer stem cells: Adding glycosylation to the equation: Drug Resist Updat, 2016; 24; 34-54
30. Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG, Cancer drug resistance: An evolving paradigm: Nat Rev Cancer, 2013; 13(10); 714-26
31. Haslehurst AM, Koti M, Dharsee M, EMT transcription factors snail and slug directly contribute to cisplatin resistance in ovarian cancer: BMC Cancer, 2012; 12; 91
32. Castellano E, Molina-Arcas M, Krygowska AA, RAS signalling through PI3-Kinase controls cell migration via modulation of Reelin expression: Nat Commun, 2016; 7; 11245
33. Du Y, Song W, Chen J, The potassium channel KCa3.1 promotes cell proliferation by activating SKP2 and metastasis through the EMT pathway in hepatocellular carcinoma: Int J Cancer, 2019; 145(2); 503-16
34. Wagner EF, Nebreda AR, Signal integration by JNK and p38 MAPK pathways in cancer development: Nat Rev Cancer, 2009; 9(8); 537-49
35. Dhillon AS, Hagan S, Rath O, Kolch W, MAP kinase signalling pathways in cancer: Oncogene, 2007; 26(22); 3279-90
36. Feng X, Liu H, Zhang Z, Annexin A2 contributes to cisplatin resistance by activation of JNK-p53 pathway in non-small cell lung cancer cells: J Exp Clin Cancer Res, 2017; 36(1); 123
37. Wei J, Li Z, Chen W, AEG-1 participates in TGF-beta1-induced EMT through p38 MAPK activation: Cell Biology Int, 2013; 37(9); 1016-21
Figures
Figure 1. Differences between A549/DDP cells and parental A549 cells. (A) A549/DDP cells show increased resistance to cisplatin compared with A549 cells, as determined by CCK-8 assays. (B) A549/DDP cells exhibit a fibroblastic morphology, while A549 cells have an epithelial-like appearance. (C–E) Real-time PCR, western blot, and immunofluorescence analyses of gene and protein expression of EMT-related markers E-cadherin and N-cadherin. (F) Migration assays revealed significant enhancement of migration ability in A549/DDP cells relative to A549 cells. (G) Protein expression of Reelin was measured by western blotting. (H) A549 cells were treated with cisplatin (1 μg/mL) for the times indicated, and the protein levels of Reelin were determined by western blotting. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8; PCR – polymerase chain reaction; EMT – epithelial-mesenchymal transition.Figure 2. Reelin enhances cisplatin resistance in lung cancer cells. (A) A549/DDP cells were transfected with Reelin siRNA and Reelin expression was examined by western blotting. (B) A549/DDP cells transfected with Reelin siRNA were treated with cisplatin at the different concentrations for 48 hours; the viability was measured by CCK-8 assays. (C) A549/DDP cells were transfected with Reelin siRNA, treated with 4 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (D) A549 cells were transfected with the Reelin expression vector, and Reelin expression was evaluated by western blotting. (E) A549 cells transfected with the Reelin expression vector were treated with cisplatin at the indicated concentrations for 48 h; cell viability was determined by CCK-8 assays. (F) A549 cells were transfected with the Reelin expression vector, treated with 2 μg/mL cisplatin for 24 hours, and stained with Annexin V-FITC and propidium iodide. Cell apoptosis was analyzed by flow cytometry. (G) H1299 and H460 cells were transfected with Reelin, and cell viability was assessed by CCK-8 assay. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8.Figure 3. Reelin promotes EMT in NSCLC cells. (A) Cell morphological observations after Reelin overexpression (A549-NC versus A549/Reelin) in A549 cells and Reelin knockdown in A549/DDP cells (A549/DDP si-NC versus A549/DDP si-Reelin). (B–D) E-cadherin and N-cadherin expressions were evaluated by western blotting and immunofluorescence in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (E, F) Cell migration was measured by Transwell migration assays. * P<0.05, ** P<0.01.Figure 4. Silencing of Snail reverses the EMT phenotype and cisplatin resistance in NSCLC cells. (A) Effects of Reelin on Snail, ZEB1, Slug, and Twist expression in A549 cells were examined by western blotting. (B) Effects of Reelin on Snail expression in A549/DDP cells were examined by western blotting. (C) Snail expression were evaluated by qRT-PCR in A549 cells transfected with the Reelin expression vector and A549/DDP cells transfected with Reelin si-RNA. (D) The expressions of E-cadherin, N-cadherin, and Snail were evaluated by western blotting after Snail knockdown in A549/Reelin cells. (E) Transwell assays revealed dramatically increased migration ability of A549/Reelin cells transfected with si-Snail relative to those transfected with si-NC. (F) A549/Reelin cells were transfected with Snail siRNA, treated with cisplatin at the indicated concentrations for 48 hours, and measured for cell viability by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8.Figure 5. Activation of the p38/GSK3β signaling pathway is involved in Reelin-induced EMT and cisplatin resistance in A549 cells. (A) A549 cells were transfected with the Reelin expression vector, and p-AKT, p-p38, p-JNK, p-ERK, β-catenin, and p-GSK-3β were evaluated by western blotting. (B) A549/DDP cells were transfected with Reelin siRNA, and p-p38 and p-GSK-3β were evaluated by western blotting. (C) A549 cells transfected with the Reelin expression vector were treated with DMSO or p38 inhibitor SB203580 (10 μM) for 48 hours, and p-GSK3β, Snail, and EMT markers were evaluated by western blotting. (D) A549 cells were treated with LiCl (40 mM) for 6 hours. The expression of Snail and p-GSK-3β were analyzed by western blotting. (E) Cell migration was measured by Transwell migration assays. (F) Cell viability was analyzed by CCK-8 assays. * P<0.05, ** P<0.01. CCK-8 – Cell Counting Kit-8. In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387