01 May 2021: Lab/In Vitro Research
Thrombin Aggravates Hypoxia/Reoxygenation Injury of Cardiomyocytes by Activating an Autophagy Pathway-Mediated by SIRT1
Xiaoning Wang1A, Yunhe Xu2BC, Lingbo Li3F, Weiwei Lu1EF*DOI: 10.12659/MSM.928480
Med Sci Monit 2021; 27:e928480






Abstract
BACKGROUND: Acute myocardial infarction is the leading cause of mortality among adults worldwide. The present study aimed to investigate the role and mechanism of thrombin and SIRT1 in hypoxia/reoxygenation (H/R) injury.
MATERIAL AND METHODS: H9c2 cardiomyocytes were used to create an H/R model to simulate in vivo ischemia/reperfusion injury. The MTT assay was used to measure cell viability, qRT-PCR was used to detect the level of SIRT1, thrombin, and PAR-1, and western blot analysis was conducted for evaluation of thrombin, PAR-1, SIRT1, LC3I, LC3II, and Beclin1. ELISA was applied for determination of IL-1β, IL-6, TNF-α, MMP-9, and ICAM-1. After the establishment of the H/R model, superoxide dismutase (SOD) activity was evaluated by the xanthine oxidase method, malondialdehyde content was detected by thiobarbituric acid assay, and reactive oxygen species generation was measured by CM-H2DCFDA.
RESULTS: The findings showed that thrombin enhanced inflammatory factor secretion and oxidative stress but inhibited cell viability in H/R-injured cardiomyocytes. We also observed that thrombin promoted autophagy in H/R-injured cardiomyocytes. In addition, thrombin enhanced the upregulation of SIRT1 expression by H/R. However, it was found that inhibition of SIRT1 could suppress the effect of thrombin on inflammatory factor secretion, oxidative stress, and cell viability. Moreover, downregulation of SIRT1 suppressed the inhibitory effect of thrombin on autophagy in H/R injury.
CONCLUSIONS: Thrombin aggravates H/R injury of cardiomyocytes by activating an autophagy pathway mediated by SIRT1. These findings might provide a potential target therapy for the treatment of ischemia/reperfusion injury in future clinical work.
Keywords: Systemic Inflammatory Response Syndrome, Thrombin, Down-Regulation, Malondialdehyde, Sirtuin 1
Background
Acute myocardial infarction (MI) is the leading cause of mortality among adults worldwide, with more than 20% of all deaths being related to cardiovascular disease in the past few decades [1,2]. Though primary percutaneous coronary intervention has been considered as a standard therapy for acute MI, myocardial ischemia/reperfusion (I/R) injury accounts for the majority of causes of heart failure [3]. I/R is a multifactorial process regulated by inflammation, metabolic factors, microvascular obstruction, and other processes and factors [4].
The serine protease thrombin is an essential component of the coagulation cascade. It is involved in endothelial cell activation, vascular biology processes, and platelet aggregation [5]. Increasing evidence has revealed that thrombin plays an important role in traumatic brain injury as well as in ischemic and hemorrhagic stroke [6]. Thrombin and thrombin receptor PAR-1 are increased in glial fibrillary acidic protein (GFAP)-positive astrocytes [7]. In addition, thrombin activity is increased in hippocampal slices after oxygen induction and glucose deprivation [8]. Moreover, thrombin has been reported to participate in the genesis of arrhythmias and activation of thrombin receptor-affected ventricular arrhythmias [9].
Autophagy refers to a process of cellular degradation, through which cellular proteins and organelles are sequestered in autophagosomes, delivered to lysosomes, and digested by lysosomal hydrolases [10]. Autophagy is an essential process in maintaining cardiomyocyte function [11]. In addition, autophagy is involved in multiple cardiovascular diseases, serving as an adaptive response or a detrimental contributor to pathogenesis [12]. Generally, autophagy plays a double role in the myocardial I/R process, in which moderate autophagy may benefit cardiomyocytes and excessive autophagy may induce harmful reactions like cell apoptosis of cardiomyocytes engaged in the I/R process [13–15]. As reported previously, moderate autophagy activation plays a cardioprotective role against I/R injury [16]. However, limited research has addressed the regulatory roles of thrombin and autophagy in myocardial ischemia-reperfusion injury. Therefore, we thought it would be highly interesting to investigate how thrombin affects autophagy in cardiomyocytes.
In the present study, the mechanism of thrombin’s effects on autophagy was deeply investigated in cardiomyocytes after hypoxia/reoxygenation injury (H/R). We demonstrated that thrombin increased H/R injury of cardiomyocytes by activating an autophagy pathway mediated by SIRT1.
Material and Methods
CELL CULTURE AND HYPOXIA/REOXYGENATION TREATMENT:
H9c2 cardiomyocytes were obtained from the American Type Culture Collection (Rockville, MD, USA). H9c2 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS) (both from Invitrogen Life Technologies, Carlsbad, CA, USA) at 37°C in a humidified incubator containing 5% CO2. The treated H9c2 cells were firstly cultured in serum-free 10% FBS DMEM with 5% CO2 at 37°C for 12 h. Then, the cells were placed in a hypoxia chamber supplemented with an anerobic pouch and incubated in 5% CO2/95% N2 at 37°C for 21 h. Subsequently, the H9c2 cells were cultured in 10% FBS DMEM in 5% CO2 at 37°C for 3 h for reoxygenation.
CELL TREATMENT AND TRANSFECTION:
For cell treatment, H/R-injured H9c2 cells were treated with either dabigatran (1 nM) or thrombin (5 U/ml) with or without dabigatran (1 nM). In addition, H/R-injured H9c2 cells were treated with thrombin (5 U/ml) with or without 3-MA. The dose of thrombin was determined according to our experimental experience and previous reference [17].
For cell transfection, H9c2 cells treated with thrombin (5 U/ml) were transfected with si-SIRT1 or si-NC (all purchased from GenePharma Co., Ltd., Shanghai, China) using Lipofectamine 2000 (Invitrogen, Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s instructions.
MTT ASSAY:
Cell viability of the H9c2 cells was detected using an MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium bromide) assay administered 48 h after transfection. Briefly, H9c2 cells were planted in 96-well plates at a density of 5×103 cells per well and incubated for 48 h. After the aforementioned treatments, 10 μl of 5 mg/mL MTT solution was added to each well and the plates were incubated for another 4 h at 37oC. After removing the MTT, the supernatant was subsequently replaced with 180 μL DMSO. The optical density (OD) value was measured at 450 nm using a microplate reader (DNM-9602; Perlong, Beijing, China).
RNA EXTRACTION AND QUANTITATIVE REAL-TIME PCR:
The expression of PAR-1, SIRT1, and thrombin was measured by quantitative Real-Time PCR (qRT-PCR). TRIZOL reagent (Invitrogen, Grand Island, NY, USA) was applied to extract total RNA. Subsequently, the High-Capacity cDNA Reverse Transcription kit (Applied Biosystems; Thermo Fisher Scientific, Inc.) was used to convert RNA into cDNA, and SYBR-Green I Real-Time PCR kit (TaKaRa Biotechnology, Dalian, China), with a Bio-Rad MiniOption thermocycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA), was used for detection. Primer sequences were as follows:
WESTERN BLOT ASSAY:
Western blotting was performed to detect the levels of thrombin, PAR-1, SIRT1, LC3I, LC3II, and Beclin1. Briefly, the total protein extracted from the sample was measured using a protein assay reagent (Bio-Rad, Hercules, CA, USA). Equal amounts of protein were separated by sodium dodecyl sulfate (10%) polyacrylamide gel electrophoresis (10% SDS-PAGE), transferred onto polyvinylidene difluoride (PVDF) membranes, and blocked with 5% nonfat milk at room temperature for 1 h. The membranes were then incubated with primary antibody overnight at 4°C: anti-thrombin (ab92621, 1 μg/ml), anti-SIRT1 (ab189494, 1/1000), anti-LC3A/B (ab128025, 2 μg/ml), anti-Beclin 1 (ab210498, 1/1000), and anti-GAPDH (ab8245, 1/500) were all purchased from Abcam (Cambridge, MA, USA), while anti-PAR1 (#MBS3008648, 1/500) was purchased from MyBioSource, Inc. (San Diego, CA, USA). After washing 3 times with tris-buffered saline with Tween, the membranes were incubated with corresponding goat anti-mouse or goat anti-rabbit secondary antibodies for 1 h at room temperature. Protein bands were analyzed using the Bio-Image Analysis System (Bio-Rad, Richmond, CA, USA). The data were normalized to b-actin, which served as an internal control.
ELISA:
The protein levels of IL-1b, IL-6, TNF-a, MMP-9, and ICAM-1 in the serum were measured using enzyme-linked immunosorbent assay (ELISA) kits. The ELISA kits used were as follows: Rat IL-1 beta ELISA Kit (ab255730), Rat IL-6 ELISA Kit (ab234570), Rat TNF alpha ELISA Kit (ab236712), Rat ICAM 1 ELISA Kit (ab100763) (all purchased from Abcam), and Rat MMP9 ELISA Kit (EK1463, Boster Biological Technology Co., Ltd). Protocols were carried out according to the manufacturer’s instructions.
MEASUREMENT OF SUPEROXIDE DISMUTASE, MALONDIALDEHYDE, AND REACTIVE OXYGEN SPECIES GENERATION:
After H/R induction, superoxide dismutase (SOD) activity was evaluated by the xanthine oxidase method, malondialdehyde (MDA) content was detected by thiobarbituric acid assay, and reactive oxygen species (ROS) generation was measured by CM-H2DCFDA (Life Technologies, Invitrogen, catalog number: C6827).
STATISTICAL ANALYSIS:
Comparison between 2 groups was performed using the
Results
THROMBIN INCREASED INFLAMMATORY FACTOR SECRETION AND OXIDATIVE STRESS IN H/R-INJURED CARDIOMYOCYTES:
To investigate the effect of thrombin on H/R-injured cardiomyocytes, inflammatory factor level, oxidative stress, and cell viability were detected. As shown in Figure 1A, MDA content and ROS level were increased following H/R treatment; however, the activity of SOD was suppressed. H/R treatment also significantly upregulated the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9 in serum (Figure 1B) but inhibited cell viability of cardiomyocytes (Figure 1C). After the H/R-injured cardiomyocytes were treated with thrombin, we observed that thrombin clearly enhanced the upregulation of MDA content, ROS level, and the level of inflammatory factors (IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9) that had been induced by H/R. Furthermore, thrombin notably promoted the inhibition effect of H/R on SOD activity and cell viability. In addition, dabigatran was able to remarkably alleviate the oxidative stress and increased inflammatory factor secretion induced by H/R and thrombin; it was also able to alleviate the effects on cell viability. All these findings suggested that thrombin increased inflammatory factor secretion and oxidative stress in H/R-injured cardiomyocytes.
THROMBIN INCREASED AUTOPHAGY IN H/R-INJURED CARDIOMYOCYTES:
Next, we studied the effect of thrombin on autophagy in H/R-injured cardiomyocytes. The western blotting result revealed that the levels of thrombin, PAR-1, SIRT1, and Beclin1, and the conversion of LC3I to LC3II, were notably increased by H/R injury (Figure 2A). Further, the effect of H/R injury on autophagy in cardiomyocytes was significantly increased by thrombin but attenuated by dabigatran. Moreover, dabigatran reversed the effect of thrombin on autophagy in H/R-injured cardiomyocytes. The qRT-PCR results also confirmed that the expression of thrombin, PAR-1, and SIRT1 were clearly upregulated by thrombin and downregulated by dabigatran in H/R-injured cardiomyocytes (Figure 2B). The above results indicated that thrombin increased autophagy in H/R-injured cardiomyocytes.
SILENCING OF SIRT1 INHIBITED THE EFFECT OF THROMBIN ON OXIDATIVE STRESS AND THE SECRETION OF INFLAMMATORY FACTORS IN H/R-INJURED CARDIOMYOCYTES:
Subsequently, we investigated the molecular mechanism whereby SIRT1 modulated the effect of thrombin on inflammatory factor secretion, oxidative stress, and cell viability in H/R-injured cardiomyocytes. We found that H/R treatment significantly increased MDA content and ROS level but inhibited SOD activity (Figure 3A). ELISA results showed that H/R also obviously upregulated serum levels of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9 (Figure 3B), while lowering cell viability in cardiomyocytes (Figure 3C). Furthermore, thrombin was also shown to notably aggravate the effect of H/R injury on cardiomyocytes. Interesting, SIRT1 knockdown or autophagy inhibitor 3-MA attenuated the level of inflammatory factors, including IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9, as well as effects on oxidative stress and cell viability, in H/R-injured cells both with and without thrombin treatment. All these findings illustrated that SIRT1 silencing inhibited thrombin-induced upregulation of the level of inflammatory factors and oxidative stress in H/R-injured cardiomyocytes.
SILENCING OF SIRT1 SUPPRESSED THE REGULATORY EFFECTS OF THROMBIN ON AUTOPHAGY IN H/R-INJURED ASTROCYTES:
We finally explored the molecular mechanism underlying the effect of SIRT1 on autophagy. Western blot (Figure 4A) revealed that the level of thrombin and PAR1 was increased by thrombin treatment, but was not affected by either si-SIRT1 or autophagy inhibitor 3-MA. Moreover, thrombin upregulated the level of autophagy factor, including the ratio of LC3II/I and Beclin1, which could be suppressed by 3-MA. SIRT1 expression was increased by thrombin and decreased by si-SIRT1. At the same time, qRT-PCR results confirmed that SIRT1 was effectively knocked down by si-SIRT1 (Figure 4B); however, the levels of thrombin and PAR1 were not obviously affected by si-SIRT1. These results indicated that silencing of SIRT1 suppressed the regulation of thrombin on autophagy in H/R-injured cardiomyocytes.
Discussion
Ischemic heart disease is characterized by restriction of blood supply to the myocardial regions, leading to infarction and tissue necrosis [18]. Moreover, myocardial infarction belongs to one of the most prevalent cardiovascular diseases worldwide [19]. In recent decades, an increasing number of studies have investigated the role of I/R in myocardial injury [20,21]; however, the molecular mechanisms involved in I/R still remain uncertain. As reported, I/R injury activates autophagy and apoptosis, leading to cell death [22]. Hence, the present study focused on the effects of thrombin on inflammatory factors and oxidative stress as well as autophagy in H/R-injured cardiomyocytes. We demonstrated, for the first time, that thrombin might promote H/R injury by activating SIRT1-induced autophagy in cardiomyocytes.
Inflammation and oxidative stress are closely associated processes that participate in various types of injury [23]. Thrombin participates in inflammation, apoptosis, and oxidative stress [24]. A previous study demonstrated that thrombin activates the NLRP3 inflammasome and induces cell apoptosis in microglia [25]. Another study suggested that thrombin induces the secretion of proinflammatory cytokines as well as enhanced inflammatory responses in the pathogenesis of atherosclerosis [26]. These results supported thrombin’s ability to activate inflammatory factors. However, as stated in another previous study, thrombin initiated proinflammatory signaling responses by activating PARs, which would serve to increase secretion of proinflammatory cytokines and chemokines, and enhance leukocyte migration [27], indicating that thrombin might play different roles in inflammation under different conditions. Our study showed that thrombin stimulated the secretion of inflammatory factors in cardiomyocytes with H/R injury, which is consistent with some of the previous findings mentioned above. However, only limited reports have illustrated the effect of thrombin on oxidative stress and autophagy. In addition, thrombin induces ROS generation, which results in oxidative damage and the death of hippocampal neurons [28]. Hu et al also demonstrated that thrombin can induce autophagy in glial cells of the rat central nervous system [29]. In addition, autophagy induced by thrombin plays a promising role in mitigating intracerebral hemorrhage [17]. These previous reports are accordant with our findings that thrombin enhanced oxidative stress and autophagy after H/R injury. Also, we showed for the first time that under H/R conditions, thrombin also promoted inflammation and oxidative stress.
SIRT1 is an NAD+-dependent protein deacetylase that is involved in various metabolic and pathological processes that protect against apoptosis, inflammation, and oxidative stress by regulating gene expression [30]. Increasing evidence indicates that SIRT1 is closely associated with inflammation and oxidative stress in the I/R process. Through targeting of SIRT1, downregulated miR-29a alleviates myocardial I/R injury through suppression of oxidative stress and NLRP3-mediated pyroptosis [31]. As reported by Zhang et al, myocardial oxidative damage and apoptosis in myocardial I/R injury of rats was attenuated by honokiol via the SIRT1-Nrf2 signaling pathway [32]. However, no study has focused on the relationship between SIRT1 and thrombin. In the present study, we demonstrated that inhibition of SIRT1 suppressed the promotion by thrombin of inflammation and oxidative stress in H/R-injured cardiomyocytes.
Moreover, SIRT1 has proved to be an important regulator of autophagy in various I/R injuries, including ischemic stroke [33], I/R liver injury [34], and myocardial I/R injury [35]. Since autophagy plays both protective and harmful roles in I/R injury, it is not surprising that SIRT1 has also been reported to improve or facilitate I/R or H/R injury. Qiu et al found that miR-204 can improve H/R injury in cardiomyocytes through inhibition of SIRT1-induced autophagy, and SIRT1 is upregulated after H/R injury [36]. However, it was also reported that under some conditions, the activation of SIRT1 might improve I/R or H/R injury [37–39]. Under these conditions, the expression of SIRT1 might be downregulated after H/R injury [40]. All these results suggest that the role of SIRT1 in I/R and H/R injury still needs more research for further characterization and confirmation. Our findings found that after H/R injury, upon thrombin treatment, the expression of SIRT1 was elevated. Silencing SIRT1, in turn, led to inhibited autophagy that was enhanced by thrombin in H/R-injured cardiomyocytes.
Conclusions
In summary, this study performed in vitro experiments designed to investigate the role of thrombin and SIRT1 in H/R injury. We demonstrated, for the first time, that thrombin affected H/R injury by activating an autophagy pathway mediated by SIRT1 in cardiomyocytes. This might provide a new direction for the development of treatments for I/R injury in the clinic.
Figures
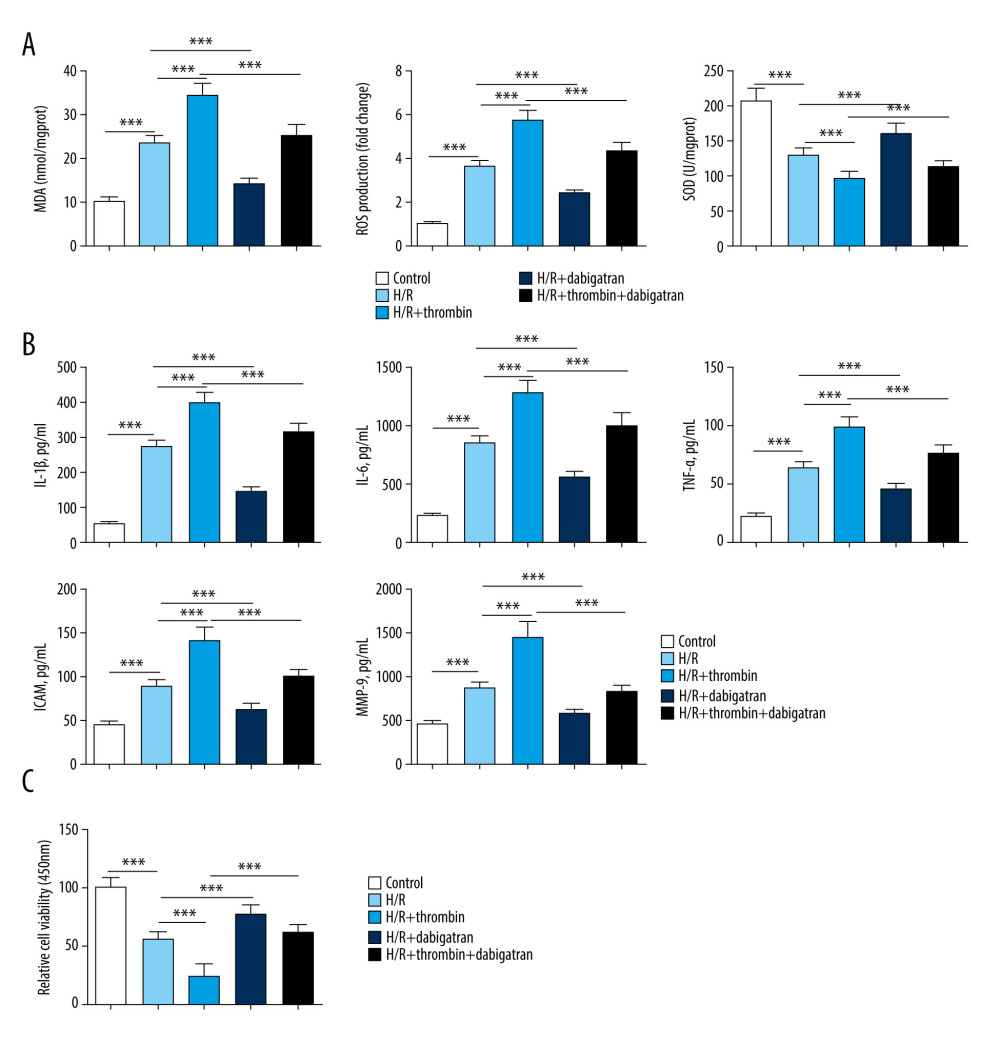 Figure 1. Induction of inflammatory factor secretion and oxidative stress by thrombin in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in the H/R model treated with thrombin, the H/R model treated with dabigatran, and the H/R model treated with a combination of thrombin and dabigatran, as well as the controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3.
Figure 1. Induction of inflammatory factor secretion and oxidative stress by thrombin in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in the H/R model treated with thrombin, the H/R model treated with dabigatran, and the H/R model treated with a combination of thrombin and dabigatran, as well as the controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3. 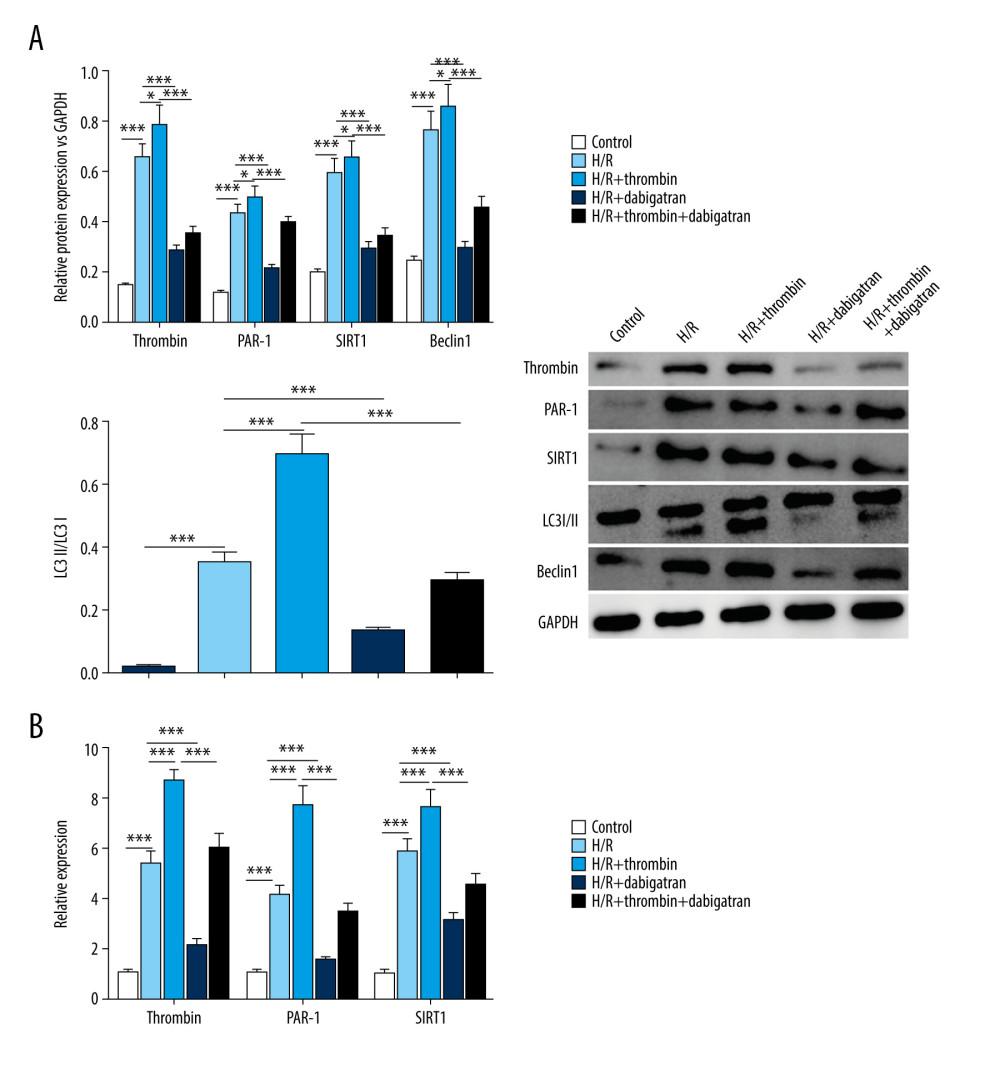 Figure 2. Induction of autophagy by thrombin in H/R-injured cardiomyocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3.
Figure 2. Induction of autophagy by thrombin in H/R-injured cardiomyocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3. 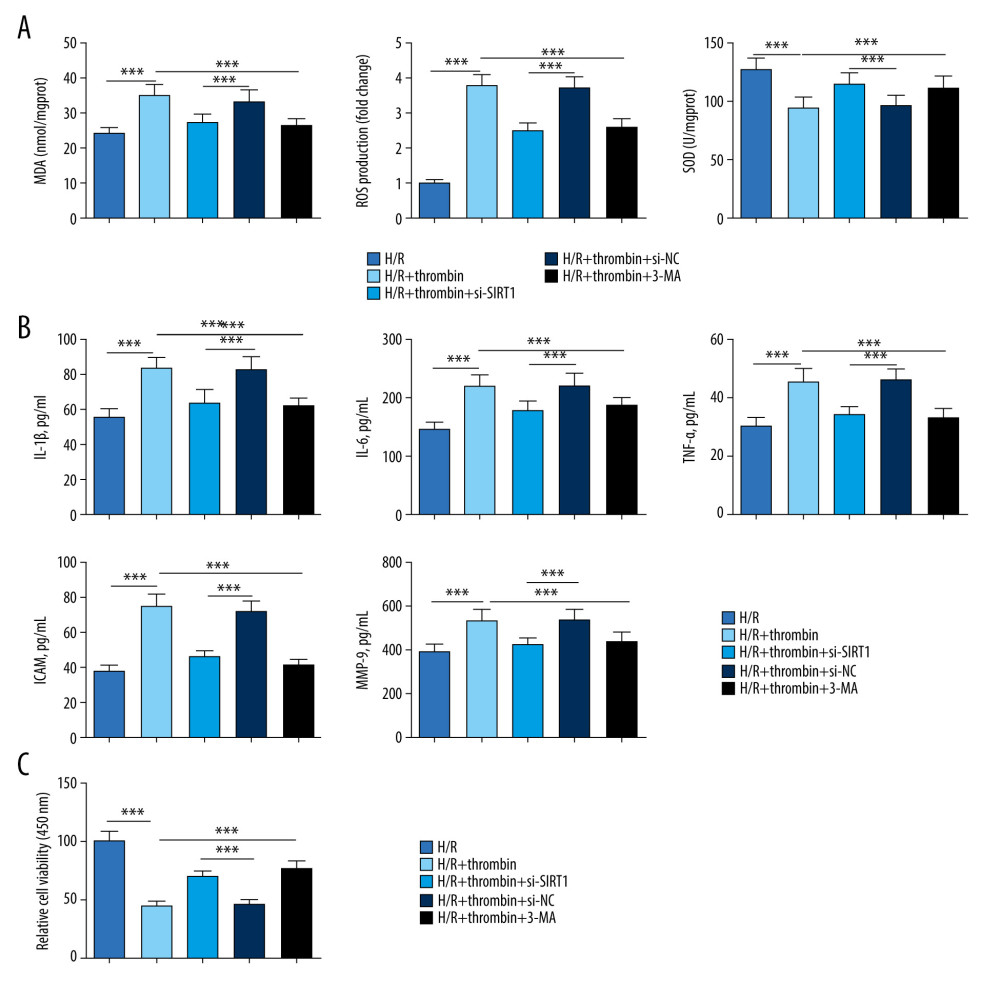 Figure 3. Silencing of SIRT1 inhibited the effect of thrombin on the secretion of inflammatory factors and oxidative stress in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in an H/R model treated with thrombin, an H/R model transfected with si-SIRT1 and dabigatran, an H/R model treated with a combination of thrombin and 3-MA, and controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9 in serum. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3.
Figure 3. Silencing of SIRT1 inhibited the effect of thrombin on the secretion of inflammatory factors and oxidative stress in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in an H/R model treated with thrombin, an H/R model transfected with si-SIRT1 and dabigatran, an H/R model treated with a combination of thrombin and 3-MA, and controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9 in serum. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3. 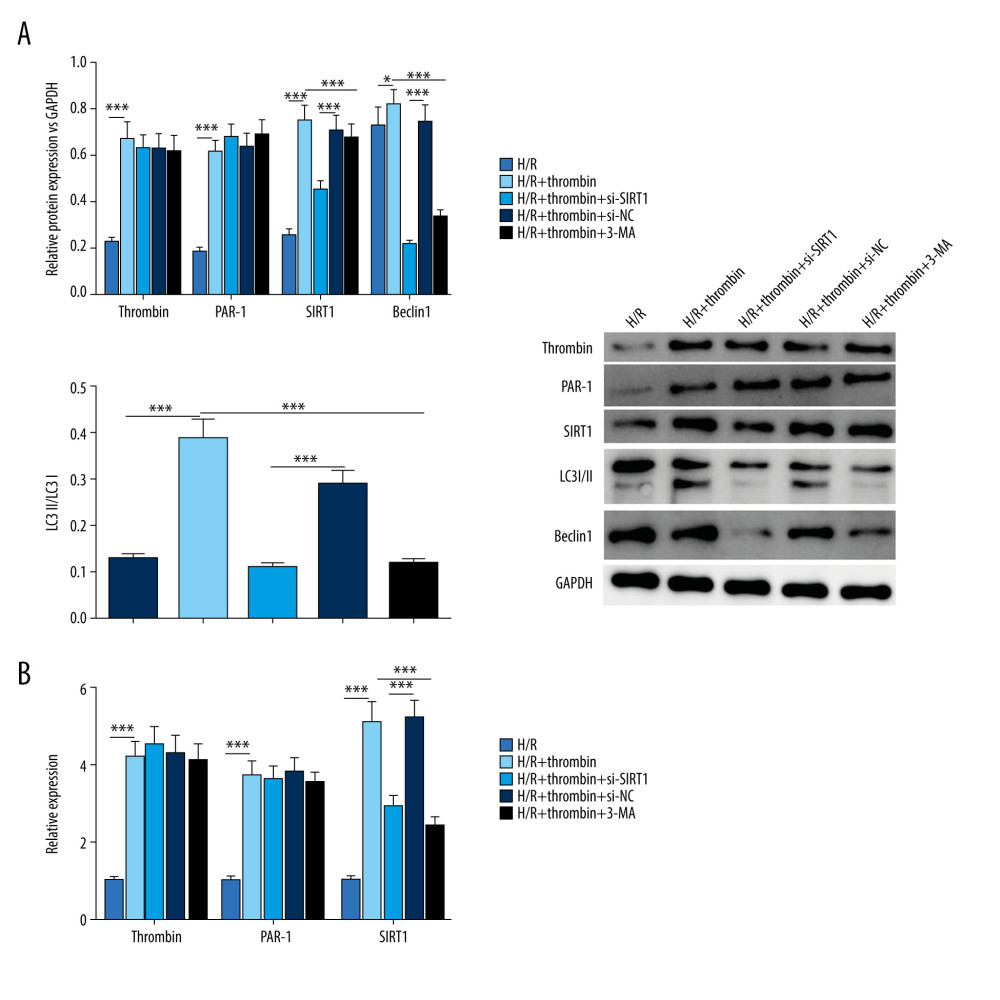 Figure 4. Silencing of SIRT1 suppressed the regulation by thrombin of autophagy in H/R-injured astrocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3.
Figure 4. Silencing of SIRT1 suppressed the regulation by thrombin of autophagy in H/R-injured astrocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3. References
1. Barnett R, Acute myocardial infarction: Lancet, 2019; 393(10191); 2580
2. Wu AH, Gornet TG, Wu VH, Early diagnosis of acute myocardial infarction by rapid analysis of creatine kinase isoenzyme-3 (CK-MM) sub-types: Clin Chem, 1987; 33(3); 358-62
3. Ibanez B, James S, Agewall S, 2017 ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation: The Task Force for the management of acute myocardial infarction in patients presenting with ST-segment elevation of the European Society of Cardiology (ESC): Eur Heart J, 2018; 39(2); 119-77
4. Davidson SM, Ferdinandy P, Andreadou I, Multitarget strategies to reduce myocardial ischemia/reperfusion injury: J Am Coll Cardiol, 2019; 73(1); 89-99
5. Negrier C, Shima M, Hoffman M, The central role of thrombin in bleeding disorders: Blood Rev, 2019; 38; 100582
6. Xing G, Deng G, Jun L, Thrombin-responsive, brain-targeting nanoparticles for improved stroke therapy: ACS Nano, 2018; 12(8); 8723-32
7. Krenzlin H, Lorenz V, Danckwardt S, The importance of thrombin in cerebral injury and disease: Int J Mol Sci, 2016; 17(1); 84
8. Stein ES, Itsekson-Hayosh Z, Aronovich A, Thrombin induces ischemic LTP (iLTP): Implications for synaptic plasticity in the acute phase of ischemic stroke: Sci Rep, 2015; 5(1); 1-7
9. Mohamed OY, Al-Masri AA, Eter EAE, Lateef R, SCH 79797, a selective PAR1 antagonist, protects against ischemia/reperfusion-induced arrhythmias in the rat hearts: Eur Rev Med Pharmacol Sci, 2016; 20(22); 4796-800
10. Levine B, Kroemer G, Biological functions of autophagy genes: A disease perspective: Cell, 2019; 176(1-2); 11-42
11. Aki T, Noritake K, Unuma K, Uemura K, Autophagic regulation of cardiomyocyte survival and heart regeneration: Autophagy in Health and Disease: Springer, 2018; 101-18
12. Ma S, Wang Y, Chen Y, Cao F, The role of the autophagy in myocardial ischemia/reperfusion injury: Biochim Biophys Acta, 2015; 1852(2); 271-76
13. Ma S, Wang Y, Chen Y, Cao F, The role of the autophagy in myocardial ischemia/reperfusion injury: Biochim Biophys Acta Mol Basis Dis, 2015; 1852(2); 271-76
14. Aghaei M, Motallebnezhad M, Ghorghanlu S, Targeting autophagy in cardiac ischemia/reperfusion injury: A novel therapeutic strategy: J Cell Physiol, 2019; 234(10); 16768-78
15. Gustafsson AsB, Gottlieb RA, Autophagy in ischemic heart disease: Circ Res, 2009; 104(2); 150-58
16. Wei C, Li H, Han L, Zhang L, Yang X, Activation of autophagy in ischemic postconditioning contributes to cardioprotective effects against ischemia/reperfusion injury in rat hearts: J Cardiovasc Pharmacol, 2013; 61(5); 416-22
17. Hu S, Xi G, Jin H, Thrombin-induced autophagy: A potential role in intracerebral hemorrhage: Brain Res, 2011; 1424; 60-66
18. Yi D, Chen G, Min Y, Bifunctional supramolecular hydrogel alleviates myocardial ischemia/reperfusion injury by inhibiting autophagy and apoptosis: J Biomed Nanotechnol, 2018; 14(8); 1458-70
19. Frank RC, Hanidziar D, Acute infection and myocardial infarction: N Engl J Med, 2019; 380(2); 171-76
20. Jaime I, Lara M, Ernesto M-M, Myocardial injury after ischemia/reperfusion is attenuated by pharmacological Galectin-3 inhibition: Sci Rep, 2019; 9(1); 1-10
21. Zhao D, Pan F, Yang S, Cardiac-derived CTRP9 protects against myocardial ischemia/reperfusion injury via calreticulin-dependent inhibition of apoptosis: Cell Death Dis, 2018; 9(7); 1-13
22. Xiaofei J, Kar-Sheng L, Qiying C, Human mesenchymal stem cell-derived exosomes reduce ischemia/reperfusion injury by the inhibitions of apoptosis and autophagy: Curr Pharm Des, 2018; 24(44); 5334-41
23. Sufang C, Xiang L, Yanling W, Ginsenoside Rb1 attenuates intestinal ischemia/reperfusion-induced inflammation and oxidative stress via activation of the PI3K/Akt/Nrf2 signaling pathway: Mol Med Rep, 2019; 19(5); 3633-41
24. Debjani T, Alma S, Yin X, Thrombin, a mediator of cerebrovascular inflammation in AD and hypoxia: Front Aging Neurosci, 2013; 5; 19
25. Ye X, Zuo D, Yu L, ROS/TXNIP pathway contributes to thrombin induced NLRP3 inflammasome activation and cell apoptosis in microglia: Biochem Biophys Res Commun, 2017; 485(2); 499-505
26. Jaberi N, Soleimani A, Pashirzad M, Role of thrombin in the pathogenesis of atherosclerosis: J Cell Biochem, 2018; 120(4); 4757-65
27. Ebrahimi S, Jaberi N, Avan A, Role of thrombin in the pathogenesis of central nervous system inflammatory diseases: J Cell Physiol, 2016; 232(3); 482-85
28. Park KW, Jin BK, Thrombin-induced oxidative stress contributes to the death of hippocampal neurons: Role of neuronal NADPH oxidase: J Neurosci, 2008; 86(5); 1053-63
29. Ding X, Hu S, Zhang Y, Wu G, Thrombin preferentially induces autophagy in glia cells in the rat central nervous system: Neurosci Lett, 2016; 630; 53-58
30. Yang C, Yang W, He Z, Kaempferol improves lung ischemia-reperfusion injury via antiinflammation and antioxidative stress regulated by SIRT1/HMGB1/NF-kB axis: Front Pharmacol, 2020; 10; 1635
31. Ding S, Liu D, Wang L, Inhibiting microRNA-29a protects myocardial ischemia-reperfusion injury by targeting SIRT1 and suppressing oxidative stress and NLRP3-mediated pyroptosis pathway: J Pharmacol Exp Ther, 2020; 372(1); 128-35
32. Zhang B, Zhai M, Li B, Honokiol ameliorates myocardial ischemia/reperfusion injury in Type 1 diabetic rats by reducing oxidative stress and apoptosis through activating the SIRT1-Nrf2 signaling pathway: Oxid Med Cell Longev, 2018; 2018; 1-16
33. Shuhu Z, Qiao B, Chu X, Kong Q, Oxymatrine attenuates cognitive deficits through SIRT1-mediated autophagy in ischemic stroke: J Neuroimmunol, 2018; 323; 136-42
34. Jing HR, Luo FW, Liu XM, Fish oil alleviates liver injury induced by intestinal ischemia/reperfusion via AMPK/SIRT-1/autophagy pathway: World J Gastroenterol, 2018; 833-43
35. Fu W, Xu H, Yu X, 20 (S)-Ginsenoside Rg2 attenuates myocardial ischemia/reperfusion injury by reducing oxidative stress and inflammation: Role of SIRT1: Res Adv, 2018; 8(42); 23947-62
36. Qiu R, Li W, Liu Y, MicroRNA-204 protects H9C2 cells against hypoxia/reoxygenation-induced injury through regulating SIRT1-mediated autophagy: Biomed Pharmacother, 2018; 100; 15-19
37. Sheng M, Lin Y, Yu W, Berberine protects against ischemia/reperfusion injury after orthotopic liver transplantation via activating Sirt1/FoxO3 alpha, induced autophagy: Biochem Biophys Res Commun, 2017; 483(2); 885-91
38. Ren Q, Hu Z, Jiang Y, SIRT1 protects against apoptosis by promoting autophagy in the oxygen glucose deprivation/reperfusion-induced injury: Front Neurol, 2019; 10; 1289
39. He Q, Li Z, Wang Y, Hou Y, Resveratrol alleviates cerebral ischemia/reperfusion injury in rats by inhibiting NLRP3 inflammasome activation through Sirt1-dependent autophagy induction: Int Immunopharmacol, 2017; 50; 208-15
40. Li D, Wang X, Huang Q, Cardioprotection of CAPE-oNO2 against myocardial ischemia/reperfusion induced ROS generation via regulating the SIRT1/eNOS/NF-kB pathway in vivo and in vitro: Redox Biol, 2018; 15; 62-73
Figures
Figure 1. Induction of inflammatory factor secretion and oxidative stress by thrombin in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in the H/R model treated with thrombin, the H/R model treated with dabigatran, and the H/R model treated with a combination of thrombin and dabigatran, as well as the controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3.Figure 2. Induction of autophagy by thrombin in H/R-injured cardiomyocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3.Figure 3. Silencing of SIRT1 inhibited the effect of thrombin on the secretion of inflammatory factors and oxidative stress in H/R-injured cardiomyocytes. (A) MDA content, ROS level, and SOD activity were detected to evaluate oxidative stress in an H/R model treated with thrombin, an H/R model transfected with si-SIRT1 and dabigatran, an H/R model treated with a combination of thrombin and 3-MA, and controls. (B) ELISA was used to determine the level of IL-1b, IL-6, TNF-a, ICAM-1, and MMP-9 in serum. (C) MTT was conducted to test cell viability. *** P<0.001 vs control group, n=3.Figure 4. Silencing of SIRT1 suppressed the regulation by thrombin of autophagy in H/R-injured astrocytes. (A) Western blotting was applied to determine the level of thrombin, PAR-1, SIRT1, Beclin1, and the conversion of LC3I to LC3II. (B) qRT-PCR was conducted to detect the expression of thrombin, PAR-1, and SIRT1. *** P<0.001 vs control group, n=3. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387