21 July 2020: Lab/In Vitro Research
Silencing of Long Non-Coding RNA (LncRNA) Non-Coding RNA Activated by DNA Damage (NORAD) Inhibits Proliferation, Invasion, Migration, and Promotes Apoptosis of Glioma Cells via Downregulating the Expression of AKR1B1
Lun Luo1ABE*, Chuan Chen1ACD, Haiyong He1DFG, Meiqin Cai1BCFG, Cong Ling1BCEDOI: 10.12659/MSM.922659
Med Sci Monit 2020; 26:e922659






Abstract
BACKGROUND: We aimed to investigate the functions of long non-coding RNA (lncRNA) non-coding RNA activated by DNA damage (NORAD) in glioma and identify the potential mechanisms.
MATERIAL AND METHODS: The expression of NORAD and AKR1B1 in human glioma cell lines were examined using reverse transcription-quantitative polymerase chain reaction (RT-qPCR). Then, cell proliferation, invasion, and migration were tested by Cell Counting Kit-8 (CCK-8), colony formation assay, Transwell, and scratch wound healing assay after NORAD silencing. Meanwhile, western blotting was utilized to measure the expression of migration-related proteins. Apoptosis of glioma cells was detected using flow cytometry and apoptosis-related proteins expression was determined. Moreover, the correlation between NORAD and AKR1B1 was verified by RNA-binding protein immunoprecipitation (RIP assay). After co-transfection with AKR1B1 overexpressed plasmid and NORAD siRNA, cell proliferation, invasion, migration, and apoptosis were examined again. Furthermore, the expression of proteins in extracellular signal-regulated kinase (ERK) signaling was tested using western blotting.
RESULTS: The results revealed that NORAD and AKR1B1 were highly expressed in glioma cells. NORAD silencing inhibited proliferation, invasion and migration but promoted apoptosis of glioma cells, accompanied by the expression changes of migration- and apoptosis-related proteins. However, after co-transfection with AKR1B1 pcDNA3.1 in NORAD silencing cells, the effects of NORAD silencing on proliferation, invasion, migration, and apoptosis were attenuated. Consistently, the expression of phosphorylated ERK (p-ERK) was decreased after NORAD silencing, which were reversed following AKR1B1 overexpression.
CONCLUSIONS: These findings demonstrated that NORAD silencing suppressed proliferation, invasion, and migration and boosted apoptosis of glioma cells via downregulating the AKR1B1 expression, which may provide a potential therapeutic target for glioma treatment.
Keywords: Aldehyde Reductase, Down-Regulation, Extracellular Signal-Regulated MAP Kinases, Gene Silencing, Glioma, Phosphorylation, Protein Binding
Background
Glioma is known as one of the most prevalent and aggressive brain tumors and it is well-known for its unfavorable prognosis, which accounts for about 80% of malignant neoplasia of the central nervous system [1,2]. Compelling evidence indicates that thousands of new patients are diagnosed with glioma each year worldwide [3]. Despite some progress in the diagnosis and treatment strategies in recent years, the overall survival rate of patients with glioma is markedly poor, which is attributed to tumor invasion and metastasis [4,5]. Therefore, a deeper understanding of the molecular mechanisms implicated in the development and progression of glioma is crucial for the prevention and treatment of glioma, which is a major task for researchers.
Long non-coding RNAs (lncRNAs) are acknowledged as one subtype of endogenous non-protein-coding transcripts, which consist of >200 nucleotides [6,7]. LncRNAs are newly identified regulators in multiple cancers. A growing body of literature has shown that aberrant levels of lncRNAs are involved in multiple biological processes of cancers through regulating the expression of genes at the transcriptional or posttranscriptional level [8,9]. It has been well documented that lncRNA non-coding RNA activated by DNA damage (NORAD) is dramatically upregulated in various malignant tumors, contributing to cancer progression [10,11]. Reports have also demonstrated that NORAD silencing can suppress proliferation, invasion, and migration but promote apoptosis in gastric cancer cells via regulation of the RhoA/ROCK1 pathway [12]. However, the functions of NORAD in glioma remain to be elucidated.
In the current study, we concentrated on the functions of NORAD on proliferation, invasion, migration, and apoptosis of glioma cells, and probed the underlying mechanisms involved in the progress of gliomas. Our findings may offer new insights into the diagnosis and strategies of glioma.
Material and Methods
CELL CULTURE:
Human glioma cell lines (GSC11, M059J, U251, and T98G) and normal human astrocytes A735 cells were obtained from the Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco; Thermo Fisher Scientific) added with 10% fetal bovine serum (FBS; Sigma-Aldrich, St Louis, MO, USA) at 37°C in a humidified incubator containing 5% CO2.
CELL TRANSFECTION:
For the transfection, U251 cells were plated in a 24-well plate. When cells reached 70% confluence, small interfering RNAs (siRNAs) against NORAD (si-NORAD-1 and si-NORAD-2), negative control for NORAD siRNA (si-NC), AKR1B1 overexpressed plasmid pcDNA3.1 (pcDNA-AKR1B1) and empty vector plasmid (pcDNA-NC) were transfected into cells using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) following manufacturer’s recommendations. The sequences used in this study were as follows:
All the aforementioned transfection complexes were purchased from GenePharma (Shanghai, China). At 48 hours post-transfection, cells were collected and the transfection efficacy was detected using reverse transcription-quantitative polymerase chain reaction (RT-qPCR) analysis or western blot analysis.
CELL PROLIFERATION ASSAY:
Cell Counting Kit-8 (CCK-8) assay was used to test proliferation of U251 cells after transfection. In brief, cells were grown in 96-well plates. At 24, 48, and 72 hours after transfection, 10 μL CCK-8 solution was appended for 2 hours of incubation. The optical density was determined at 450 nm using a microplate reader.
TRANSWELL ASSAY:
The ability of cell invasion was examined utilizing Transwell assay. 24-well Transwell plates (Corning, Inc.) with 8-μm pore inserts coated with Matrigel (BD Biosciences) was used in this experiment. U251 cells were re-suspended in 200 μL serum-free DMEM (5×104 cells/well) and put into the upper compartment. Then, 600 μL DMEM containing 10% FBS was added to the lower chamber. Following incubation for 24 hours, a cotton-tipped swab was employed to remove cells remaining on the upper surface of the filter. Cells transferred to the basolateral chamber were fixed with 4% paraformaldehyde and stained with 1% crystal violet solution. The invasive cells in 5 random fields (magnification 200x) were counted under an inverted microscope (Olympus Corporation, Tokyo, Japan).
SCRATCH WOUND HEALING ASSAY:
The migratory ability of U251 cells was determined using scratch wound healing assay. Briefly, cells were plated in a 6-well plate and cultured at 37°C in a humidified incubator until 80% confluence. Subsequently, the cells had been serum-starved in serum-free medium for 24 hours. A 200 μL of pipette tip was then applied to create a wound on the surface of monolayers. The cells were washed twice with phosphate-buffered saline (PBS) in order to remove debris and then maintained in serum-free DMEM for 48 hours. Wound healing was observed using microscopy (magnification 100×; Olympus Corporation, Tokyo, Japan), the percentage of which was calculated using ImageJ software (NIH, MA, USA). The average distance of cells migrated into wound surface was detected 48 hours later under an inverted microscope.
CELL APOPTOSIS ASSAY:
Transfected U251 cells were collected and then resuspended in the 1x binding buffer to a concentration of 1×104 cells/well. An Annexin V-FITC Staining Cell Apoptosis Detection kit (KeyGEN BioTECH, China) was adopted for assessing cell apoptosis following manufacturer’s recommendations. Briefly, cells were stained with Annexin V-FITC and propidium iodide (PI) and incubated in the dark. Cell apoptosis was analyzed with FlowJo software (Becton-Dickinson-San Jose, CA, USA).
RT-QPCR:
Total RNA extraction was conducted using TRIzol® reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Then, cDNA was synthesized by use of a Reverse Transcription kit (Beijing TransGen Biotech, Beijing, China) in accordance with the manufacturer’s guidelines. Samples were prepared for PCR using iTaq™ Universal SYBR® Green Supermix (Bio-Rad Laboratories, Inc.) on an ABI 7500 instrument (Applied Biosystems; Foster City, CA, USA). GAPDH was considered as internal reference gene. Relative expression was normalized against GAPDH and calculated based on the 2−ΔΔCq method. The qPCR primers were as follows:
NORAD forward, 5′-AAGCTGCTCTCAACTCCACC-3′ and
RNA IMMUNOPRECIPITATION (RIP) ASSAY:
The starBase database predicated that AKR1B1 could target NORAD. Therefore, RIP assay was employed to verify the binding between them. The experiment was performed using a RNA-Binding Protein Immunoprecipitation kit (Sigma-Aldrich; Merck KGaA). In brief, cells were lysed in lysis buffer and the cell lysates were incubated with magnetic beads conjugated to human anti-argonaute2 (Ago2) antibody (Abcam) or anti-IgG (negative control, EMD Millipore). The enrichment of NORAD and AKR1B1 was tested using RT-qPCR.
WESTERN BLOT ANALYSIS:
Cellular total protein was extracted from U251 cells using a protein lysis buffer in the presence of a protease inhibitor cocktail (Beyotime, Shanghai, China). A BCA Protein Quantitation kit (Beyotime, Shanghai, China) was executed for detecting the concentration of protein in each group. Equal amounts (40 μg) of protein samples were resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membrane (Millipore, USA). Subsequently, the membranes were blocked in 5% skimmed milk and then incubated with primary antibodies. After washing, proteins on the PVDF membranes were incubated with horseradish peroxidase-conjugated secondary antibodies and visualized using a chemiluminescence kit (Pierce Biotechnology; Thermo Fisher Scientific, Inc). Intensity of bands were quantified by using ImageJ software (NIH, MA, USA). GAPDH was used as internal control.
STATISTICAL ANALYSIS:
The data in the present study were presented as mean±standard deviation (SD). All data were compared using Student’s
Results
THE EXPRESSION OF NORAD WAS HIGHLY EXPRESSED IN GLIOMA CELL LINES:
To explore the function of NORAD in glioma, the level of NORAD in several glioma cell lines (GSC11, M059J, U251, and T98G) were tested using RT-qPCR. Results suggested that NORAD was highly expressed in glioma cell lines relative to normal human astrocytes A735 cells, with the highest NORAD level in U251 cells (Figure 1A). Therefore, the U251 cell line was used to perform the following experiments in the present study. Then, si-NORAD-1 or si-NORAD-2 were transfected into U251 cells and the transfection efficiency was affirmed using RT-qPCR (Figure 1B). Cells transfected with si-NORAD-2 were selected for further assays due to its higher inference efficacy. The results indicated that NORAD might possess a prominent role in glioma.
NORAD SILENCING HAMPERED PROLIFERATION, INVASION, AND MIGRATION OF GLIOMA CELLS:
The roles of NORAD silencing on proliferation, invasion, and migration of glioma cells were investigated subsequently. The results of CCK-8 (Figure 2A) and colony formation assay (Figure 2B) demonstrated that proliferation was obviously suppressed in si-NORAD-2 group compared with the si-NC group. In addition, the ability of cell invasion and migration were assessed by Transwell assay and scratch wound healing assay, respectively. As exhibited in Figure 2C and 2D, transfection with si-NORAD-2 restrained glioma cell invasion in contrast with transfection with si-NC. The changes of cell migratory ability (Figure 2E, 2F) and expression of migration-related genes MMP2, MMP9, and vimentin (Figure 3A, 3B) had the same trend with cell invasion. Taken together, these results implicated that NORAD silencing suppressed proliferation, invasion, and migration of glioma cells.
NORAD SILENCING PROMOTED APOPTOSIS OF GLIOMA CELLS:
To confirm the effect of NORAD silencing on apoptosis of glioma cells, flow cytometry analysis was performed in the study. Significant increase of apoptotic rate was observed in U251 cells after transfection with si-NORAD-2 relative to the si-NC group (Figure 4A, 4B). Meanwhile, the expression of apoptosis-related proteins was measured using western blotting. Results demonstrated that NORAD inhibition lowered the expression of Bcl-2, accompanied by increased expression of Bax, caspase-3, and caspase-9 (Figure 4C), hinting that cell apoptosis was accelerated after NORAD was silenced. These observations revealed that NORAD downregulation activated apoptosis of glioma cells.
AKR1B1 COULD BIND TO NORAD:
To further explore the NORAD-mediated molecular mechanism, the starBase database was employed to predict the potential targets of NORAD, and AKR1B1 was predicted as a target of NORAD, which was verified by RIP assay (Figure 5A). Then, the protein and messenger RNA (mRNA) expression of AKR1B1 in glioma cell lines was evaluated using western blotting and RT-qPCR, respectively. As presented in Figure 5B and 5C, the protein and mRNA levels of AKR1B1 were notably upregulated in glioma cell lines, especially in U251 cells. Also, obviously downregulated expression of AKR1B1 protein and mRNA was observed after NORAD silencing (Figure 5D, 5E). Overall, these data suggested that AKR1B1 could bind to NORAD.
AKR1B1 OVEREXPRESSION REVERSED THE INHIBITORY EFFECTS OF NORAD DOWNREGULATION ON PROLIFERATION, INVASION, AND MIGRATION OF GLIOMA CELLS:
To confirm the molecular mechanism of NORAD and AKR1B1 in glioma, AKR1B1 overexpressed plasmid pcDNA3.1 was transfected into U251 cells. As shown in Figure 6A, the expression of AKR1B1 was significantly upregulated in pcDNA-AKR1B1 relative to the pcDNA-NC group. Results of CCK-8 assay (Figure 6B) and colony formation assay (Figure 6C) demonstrated that AKR1B1 overexpression reversed the inhibition of NORAD silencing on proliferation of U251 cells. Concurrently, compared with the si-NORAD-2+pcDNA-NC group, the invasiveness and migration of U251 cells were increased after co-transfection of si-NORAD-2 and pcDNA-AKR1B1 (Figure 6D–6G). Moreover, changes of MMP2, MMP9, and vimentin expression presented the same results with the aforementioned findings (Figure 7A, 7B). These findings indicated that AKR1B1 overexpression restored the effects of NORAD silence on proliferation, invasion, and migration of glioma cells.
AKR1B1 OVEREXPRESSION ATTENUATED THE PROMOTIVE EFFECT OF NORAD DOWNREGULATION ON APOPTOSIS OF GLIOMA CELLS:
Apoptosis of glioma cells was tested using flow cytometry analysis. As determined in Figure 8A and 8B, the rate of apoptotic cells was dramatically decreased after AKR1B1 overexpression in U251 cells with NORAD silencing. Meanwhile, co-transfection with si-NORAD-2 and pcDNA-AKR1B1 upregulated the expression of Bcl-2 as well as downregulated the expression of Bax, caspase-3, and caspase-9 in comparison to co-transfection with si-NORAD-2 and pcDNA-NC (Figure 8C). Taken together, the data suggested that AKR1B1 overexpression relieved the promotion of NORAD silencing on apoptosis of glioma cells.
NORAD COULD REGULATE EXTRACELLULAR SIGNAL-REGULATED KINASE (ERK) SIGNALING VIA TARGETING AKR1B1:
To further explicit the molecular mechanism of NORAD and AKR1B1 in glioma, the expression of extracellular signal-regulated kinase (ERK) signaling was examined using western blotting. Result in Figure 9 shows that the expression of p-ERK was remarkably reduced after NORAD was decreased, which was abolished following co-transfection with si-NORAD-2 and pcDNA-AKR1B1. Overall, the finding illustrated that NORAD could regulate ERK signaling via targeting AKR1B1 in glioma.
Discussion
A growing body of evidence suggests that an increasing number of lncRNAs have been proven to be implicated in the progression of glioma [13,14]. The findings of this current study delineated that lncRNA NORAD was highly expressed in glioma cells, and silencing of NORAD suppressed the proliferation, invasion, and migration while promoting the apoptosis of glioma cells through downregulating the expression of AKR1B1.
Previous studies in the last several decades have indicated that NORAD serves a pivotal regulator in the occurrence and progression of a variety of malignancies, including gastric cancer, prostate cancer, and osteosarcoma [10,15,16]. Overexpression of NORAD is closely related to poor prognosis in various patients with cancer, such as esophageal squamous cell carcinoma, gastric cancer, and ovarian cancer [17–19]. In the present study, highly expressed NORAD was identified in several glioma cell lines, including U251. We transfected NORAD siRNA into U251 cells to successfully knock-down NORAD expression for exploring the function of NORAD in glioma. Mounting evidence has highlighted the importance of NORAD in proliferation, invasion, migration, and apoptosis of cancer cells [12,20]. Our results consistently found that silencing of NORAD caused significant declines in proliferation, invasion, migration, and resistance to apoptosis, accompanied by inhibition of migration-related proteins MMP2, MMP9, and vimentin, downregulation of anti-apoptosis protein Bcl-2 and upregulation of pro-apoptosis proteins Bax, caspase-3, and caspase-9. Our study, for the first time, uncovered the aberrant expression and regulatory function of NORAD in glioma, suggesting a novel diagnostic and therapeutic target for glioma.
To identify the potential mechanism underlying NORAD in glioma, we searched on starBase database and found that NORAD could target AKR1B1, which was further verified by RIP assay. It has been reported that overexpression of AKR1B1 is highly correlated with basal-like breast cancer and predicts poor prognosis of breast cancer patients [21]. It is noteworthy that upregulated expression of AKR1B1 can promote proliferation and suppress apoptosis of pancreatic cancer cells via the ERK1/2 pathway [22]. Moreover, numerous studies have unveiled that activation of ERK signaling promotes the progression of glioma [23,24]. Therefore, we deduced that NORAD might upregulate the expression of AKR1B1 to activate the ERK pathway, thus developing its carcinogenic role in glioma cells. In the current paper, obviously increased expression of AKR1B1 was observed in glioma cells. Importantly, AKR1B1 overexpression reversed the effects of NORAD silencing on proliferation, invasion, migration, and apoptosis of glioma cells, coupled with changes of migration- and apoptosis-related proteins. Besides, silencing of NORAD remarkably restrained the expression of phosphorylated ERK (p-ERK), which was restored by AKR1B1 overexpression.
Conclusions
Taken together, these findings elucidated that lncRNA NORAD was upregulated in glioma. NORAD silencing suppressed cellular proliferation, invasion, migration, and promoted apoptosis, whereas these impacts were rescued after AKR1B1 overexpression via activation of the ERK signaling. The present findings demonstrated that NORAD might be a novel therapeutic target for the treatment of glioma. However, the usage of a single glioma cell line and lack of studies
Figures
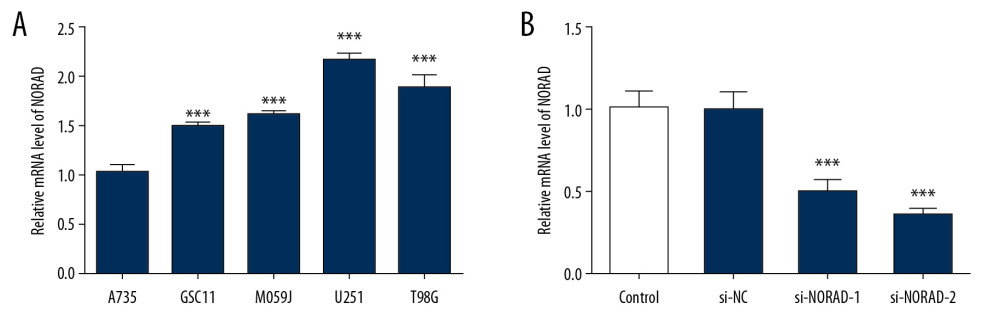 Figure 1. NORAD was highly expressed in glioma cell lines. (A) The expression of NORAD in glioma cell lines (GSC11, M059J, U251, and T98G) and normal human astrocytes A735 cells was detected using RT-qPCR. Three independent experiments were carried out (n=3). *** P<0.001 vs. A735. (B) The expression of NORAD was tested using RT-qPCR after transfection with si-NORAD-1 or si-NORAD-2. Three independent experiments were carried out (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD.
Figure 1. NORAD was highly expressed in glioma cell lines. (A) The expression of NORAD in glioma cell lines (GSC11, M059J, U251, and T98G) and normal human astrocytes A735 cells was detected using RT-qPCR. Three independent experiments were carried out (n=3). *** P<0.001 vs. A735. (B) The expression of NORAD was tested using RT-qPCR after transfection with si-NORAD-1 or si-NORAD-2. Three independent experiments were carried out (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD. 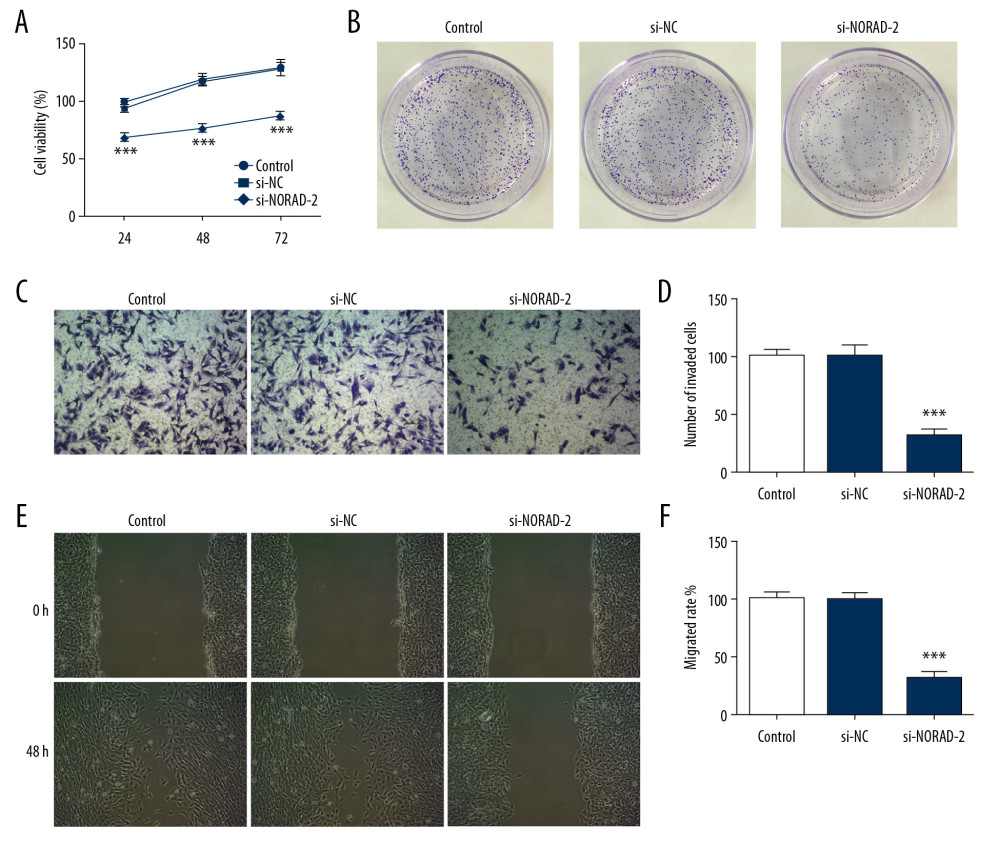 Figure 2. NORAD silencing inhibited proliferation, invasion, and migration of glioma cells. Cell proliferation was examined using (A) CCK-8 and (B) colony formation assay. (C) The invasion ability of U251 cells was evaluated using Transwell assay. (magnification 200×). (D) Quantification of cell invasion. (E) Representative images and (F) relative quantification of cell migration, as measured using wound healing assay. (magnification 100×). All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA.
Figure 2. NORAD silencing inhibited proliferation, invasion, and migration of glioma cells. Cell proliferation was examined using (A) CCK-8 and (B) colony formation assay. (C) The invasion ability of U251 cells was evaluated using Transwell assay. (magnification 200×). (D) Quantification of cell invasion. (E) Representative images and (F) relative quantification of cell migration, as measured using wound healing assay. (magnification 100×). All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA. 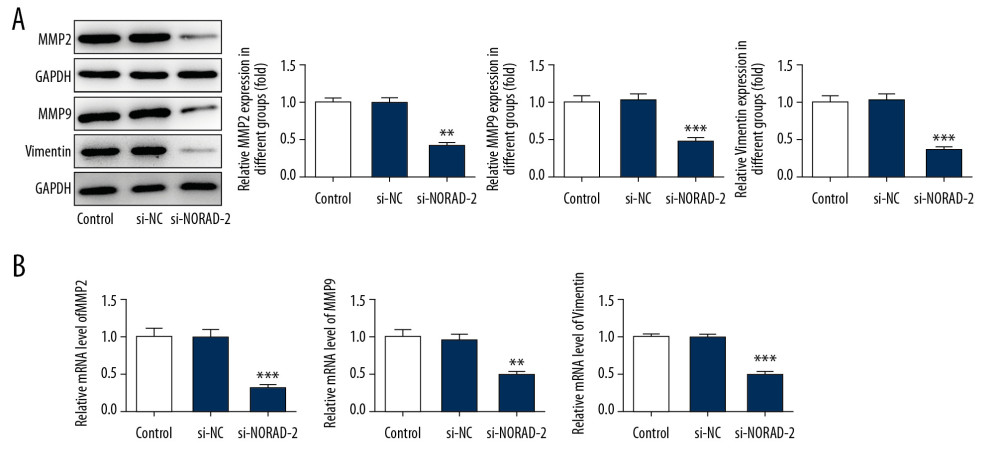 Figure 3. NORAD silencing reduced the expression of migration-related genes in glioma cells. The MMP2, MMP9, and vimentin (A) protein and (B) mRNA expression was detected using western blotting and RT-qPCR, respectively. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA.
Figure 3. NORAD silencing reduced the expression of migration-related genes in glioma cells. The MMP2, MMP9, and vimentin (A) protein and (B) mRNA expression was detected using western blotting and RT-qPCR, respectively. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA. 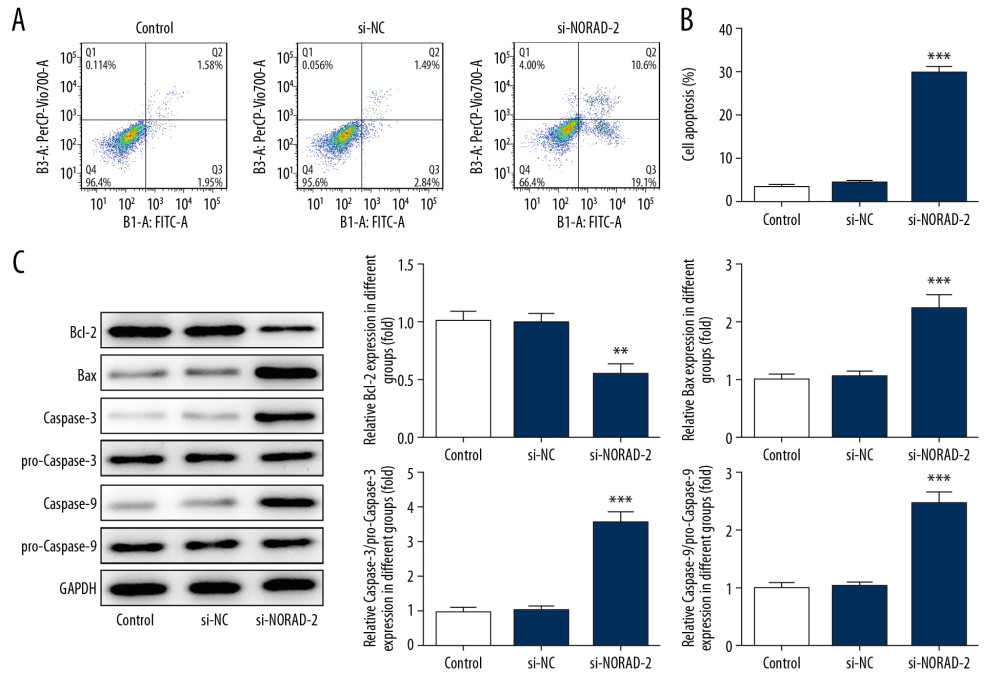 Figure 4. (A–C) NORAD silencing promoted apoptosis of glioma cells. (A) Flow cytometry analysis was used to assess the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – si-NC, negative control for NORAD siRNA.
Figure 4. (A–C) NORAD silencing promoted apoptosis of glioma cells. (A) Flow cytometry analysis was used to assess the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – si-NC, negative control for NORAD siRNA. 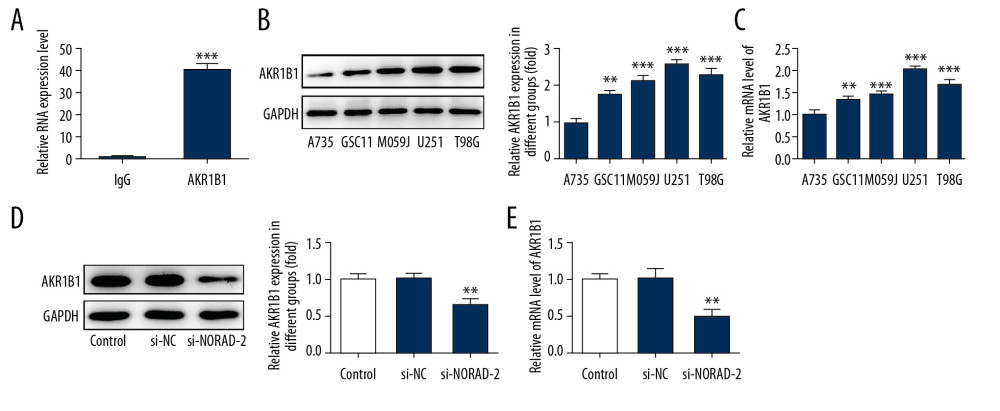 Figure 5. AKR1B1 interacted with NORAD. (A) The interplay between NORAD and AKR1B1 was detected by RIP assay. The expression of AKR1B1 in glioma cell lines (GSC11, M059J, U251, and T98G) was measured by (B) western blotting and (C) RT-qPCR, respectively. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 versus A735. The level of AKR1B1 in U251 cells was examined using (D) western blotting and (E) RT-qPCR after transfection with si-NORAD-2. All experiments were repeated 3 times independently (n=3). ** P<0.01 versus si-NC. NORAD – noncoding RNA activated by DNA damage; RIP – RNA-binding protein immunoprecipitation; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD; si-NC – negative control for NORAD siRNA.
Figure 5. AKR1B1 interacted with NORAD. (A) The interplay between NORAD and AKR1B1 was detected by RIP assay. The expression of AKR1B1 in glioma cell lines (GSC11, M059J, U251, and T98G) was measured by (B) western blotting and (C) RT-qPCR, respectively. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 versus A735. The level of AKR1B1 in U251 cells was examined using (D) western blotting and (E) RT-qPCR after transfection with si-NORAD-2. All experiments were repeated 3 times independently (n=3). ** P<0.01 versus si-NC. NORAD – noncoding RNA activated by DNA damage; RIP – RNA-binding protein immunoprecipitation; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD; si-NC – negative control for NORAD siRNA. 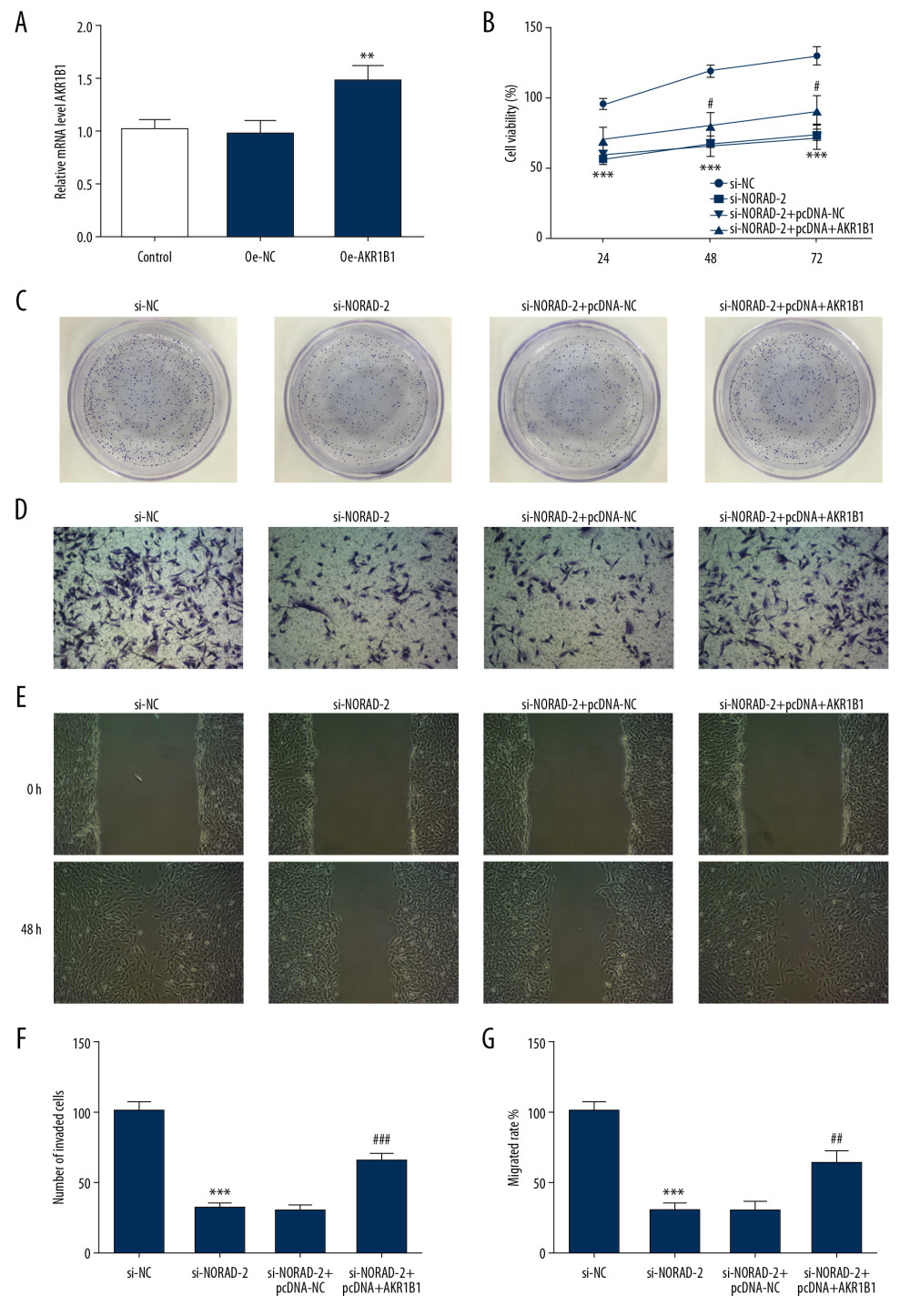 Figure 6. AKR1B1 overexpression reversed the effects of NORAD silencing on proliferation, invasion, and migration of glioma cells. (A) The expression of AKR1B1 was evaluated using RT-qPCR after transfection with AKR1B1 pcDNA3.1. Three independent experiments were carried out (n=3). ** P<0.01 versus pcDNA-NC. (B) CCK-8 assay and (C) colony formation assay tested the proliferation of U251 cells. (D) Invasion of U251 cells was evaluated using Transwell assay (magnification 200×). (E) Representative images of cell migration (magnification 100×). (F) Quantification of cell invasion. (G) Relative quantification, as examined using wound healing assay. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05, ## P<0.01, ### P<0.001 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; pcDNA-AKR1B1 – AKR1B1 overexpressed plasmid pcDNA3.1; pcDNA-NC – empty vector plasmid; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD.
Figure 6. AKR1B1 overexpression reversed the effects of NORAD silencing on proliferation, invasion, and migration of glioma cells. (A) The expression of AKR1B1 was evaluated using RT-qPCR after transfection with AKR1B1 pcDNA3.1. Three independent experiments were carried out (n=3). ** P<0.01 versus pcDNA-NC. (B) CCK-8 assay and (C) colony formation assay tested the proliferation of U251 cells. (D) Invasion of U251 cells was evaluated using Transwell assay (magnification 200×). (E) Representative images of cell migration (magnification 100×). (F) Quantification of cell invasion. (G) Relative quantification, as examined using wound healing assay. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05, ## P<0.01, ### P<0.001 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; pcDNA-AKR1B1 – AKR1B1 overexpressed plasmid pcDNA3.1; pcDNA-NC – empty vector plasmid; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD. 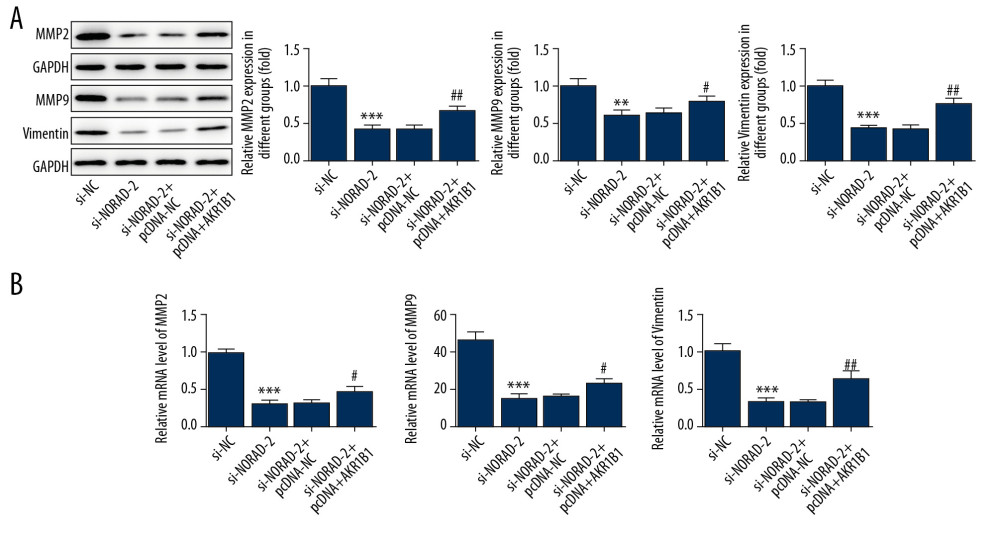 Figure 7. NORAD silencing decreased the expression of migration-related genes in glioma cells. The (A) proteins and (B) mRNA expression of MMP2, MMP9, and vimentin was detected using western blotting and RT-qPCR, respectively. All experiments were repeated three times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC; # P<0.05, ## P<0.01 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid.
Figure 7. NORAD silencing decreased the expression of migration-related genes in glioma cells. The (A) proteins and (B) mRNA expression of MMP2, MMP9, and vimentin was detected using western blotting and RT-qPCR, respectively. All experiments were repeated three times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC; # P<0.05, ## P<0.01 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid. 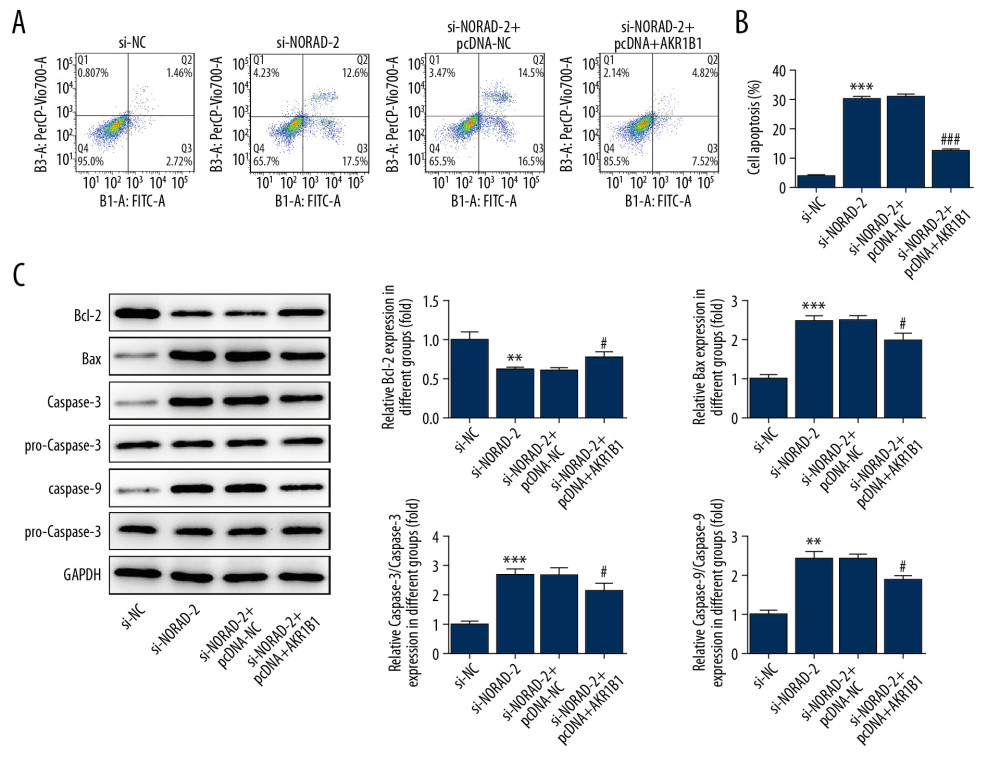 Figure 8. (A–C) AKR1B1 overexpression attenuated the effects of NORAD silencing on apoptosis of glioma cells. (A) Flow cytometry analysis detected the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 vs. si-NC; # P<0.05, ### P<0.001 versus si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid.
Figure 8. (A–C) AKR1B1 overexpression attenuated the effects of NORAD silencing on apoptosis of glioma cells. (A) Flow cytometry analysis detected the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 vs. si-NC; # P<0.05, ### P<0.001 versus si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid. 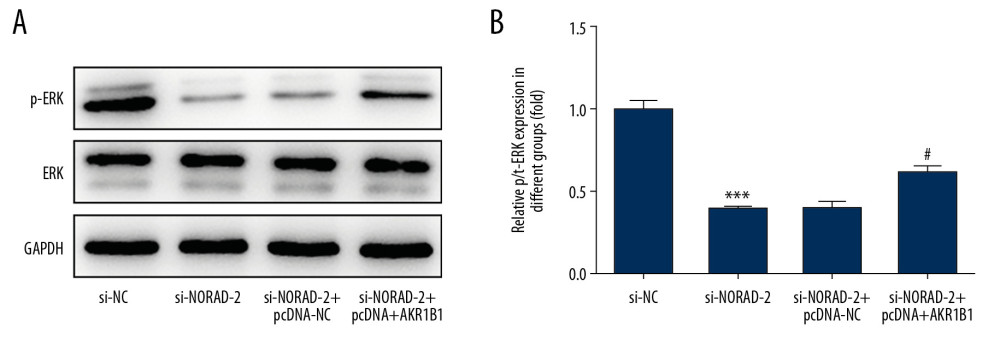 Figure 9. (A, B) NORAD could regulate ERK signaling via targeting AKR1B1. The expression of p-ERK was measured using western blotting. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; ERK – extracellular signal-regulated kinase; p-ERK – phosphorylated ERK; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid.
Figure 9. (A, B) NORAD could regulate ERK signaling via targeting AKR1B1. The expression of p-ERK was measured using western blotting. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; ERK – extracellular signal-regulated kinase; p-ERK – phosphorylated ERK; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid. References
1. Gagliardi F, Narayanan A, Reni M, The role of CXCR4 in highly malignant human gliomas biology: Current knowledge and future directions: Glia, 2014; 62; 1015-23
2. Zhou S, Ding F, Gu X, Non-coding RNAs as emerging regulators of neural injury responses and regeneration: Neurosci Bull, 2016; 32; 253-64
3. Rao AD, Rashid AS, Chen Q, Reirradiation for recurrent pediatric central nervous system malignancies: A multi-institutional review: Int J Radiat Oncol Biol Phys, 2017; 99; 634-41
4. Liu L, Cui S, Wan T, Long non-coding RNA HOTAIR acts as a competing endogenous RNA to promote glioma progression by sponging miR-126-5p: J Cell Physiol, 2018; 233; 6822-31
5. Sathornsumetee S, Rich JN, New treatment strategies for malignant gliomas: Expert Rev Anticancer Ther, 2006; 6; 1087-104
6. Chen LL, Zhao JC, Functional analysis of long noncoding RNAs in development and disease: Adv Exp Med Biol, 2014; 825; 129-58
7. Iyer MK, Niknafs YS, Malik R, The landscape of long noncoding RNAs in the human transcriptome: Nat Genet, 2015; 47; 199-208
8. Huarte M, The emerging role of lncRNAs in cancer: Nat Med, 2015; 21; 1253-61
9. Prensner JR, Chinnaiyan AM, The emergence of lncRNAs in cancer biology: Cancer Discov, 2011; 1; 391-407
10. Wang XM, Zou JL, Chen HJ, Long noncoding RNA NORAD regulates cancer cell proliferation and migration in human osteosarcoma by endogenously competing with miR-199a-3p: IUBMB Life, 2019; 71; 1482-91
11. Chen TY, Qin SY, Gu YN, Long non-coding RNA NORAD promotes the occurrence and development of non-small cell lung cancer by adsorbing MiR-656-3p: Mol Genet Genom Med, 2109; 7; 11
12. Yu SY, Peng H, Zhu Q, Silencing the long noncoding RNA NORAD inhibits gastric cancer cell proliferation and invasion by the RhoA/ROCK1 pathway: Eur Rev Med Pharmacol Sci, 2019; 23; 3760-70
13. Zhang D, Zhou H, Liu J, Mao J, Long noncoding RNA ASB16-AS1 promotes proliferation, migration, and invasion in glioma cells: Biomed Res Int, 2019; 2019 5437531
14. Li X, Zhang H, Wu X, Long noncoding RNA DLX6-AS1 accelerates the glioma carcinogenesis by competing endogenous sponging miR-197-5p to relieve E2F1: Gene, 2019; 686; 1-7
15. Tao W, Li Y, Zhu M, LncRNA NORAD promotes proliferation and inhibits apoptosis of gastric cancer by regulating miR-214/Akt/mTOR axis: Onco Targets Ther, 2019; 12; 8841-51
16. Zhang H, Guo H, Long non-coding RNA NORAD induces cell proliferation and migration in prostate cancer: J Int Med Res, 2019; 47; 3898-904
17. Wu X, Lim ZF, Li Z, NORAD expression is associated with adverse prognosis in esophageal squamous cell carcinoma: Oncol Res Treat, 2017; 40; 370-74
18. Miao Z, Guo X, Tian L, The long noncoding RNA NORAD promotes the growth of gastric cancer cells by sponging miR-608: Gene, 2019; 687; 116-24
19. Xu C, Zhu LX, Sun DM, Regulatory mechanism of lncRNA NORAD on proliferation and invasion of ovarian cancer cells through miR-199a-3p: Eur Rev Med Pharmacol Sci, 2020; 24; 1672-81
20. Wang LL, Du LT, Duan WL, Overexpression of long noncoding RNA NORAD in colorectal cancer associates with tumor progression: OncoTargets Ther, 2018; 11; 6757-66
21. Wu XB, Li XL, Fu Q, AKR1B1 promotes basal-like breast cancer progression by a positive feedback loop that activates the EMT program: J Exp Med, 2017; 214; 1065-79
22. Xiao MB, Jin DD, Jiao YJ, Beta 2-AR regulates the expression of AKR1B1 in human pancreatic cancer cells and promotes their proliferation via the ERK1/2 pathway: Mol Biol Rep, 2018; 45; 1863-71
23. Jin T, Li DB, Yang T, PTPN1 promotes the progression of glioma by activating the MAPK/ERK and PI3K/AKT pathways and is associated with poor patient survival: Oncol Rep, 2019; 42; 717-25
24. Mu PF, Liu K, Lin QY, Sirtuin 7 promotes glioma proliferation and invasion through activation of the ERK/STAT3 signaling pathway: Oncol Lett, 2019; 17; 1445-52
Figures
Figure 1. NORAD was highly expressed in glioma cell lines. (A) The expression of NORAD in glioma cell lines (GSC11, M059J, U251, and T98G) and normal human astrocytes A735 cells was detected using RT-qPCR. Three independent experiments were carried out (n=3). *** P<0.001 vs. A735. (B) The expression of NORAD was tested using RT-qPCR after transfection with si-NORAD-1 or si-NORAD-2. Three independent experiments were carried out (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD.Figure 2. NORAD silencing inhibited proliferation, invasion, and migration of glioma cells. Cell proliferation was examined using (A) CCK-8 and (B) colony formation assay. (C) The invasion ability of U251 cells was evaluated using Transwell assay. (magnification 200×). (D) Quantification of cell invasion. (E) Representative images and (F) relative quantification of cell migration, as measured using wound healing assay. (magnification 100×). All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC. NORAD – noncoding RNA activated by DNA damage; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA.Figure 3. NORAD silencing reduced the expression of migration-related genes in glioma cells. The MMP2, MMP9, and vimentin (A) protein and (B) mRNA expression was detected using western blotting and RT-qPCR, respectively. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA.Figure 4. (A–C) NORAD silencing promoted apoptosis of glioma cells. (A) Flow cytometry analysis was used to assess the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. All experiments were repeated 3 times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – si-NC, negative control for NORAD siRNA.Figure 5. AKR1B1 interacted with NORAD. (A) The interplay between NORAD and AKR1B1 was detected by RIP assay. The expression of AKR1B1 in glioma cell lines (GSC11, M059J, U251, and T98G) was measured by (B) western blotting and (C) RT-qPCR, respectively. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 versus A735. The level of AKR1B1 in U251 cells was examined using (D) western blotting and (E) RT-qPCR after transfection with si-NORAD-2. All experiments were repeated 3 times independently (n=3). ** P<0.01 versus si-NC. NORAD – noncoding RNA activated by DNA damage; RIP – RNA-binding protein immunoprecipitation; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NORAD – small interfering RNAs (siRNAs) against NORAD; si-NC – negative control for NORAD siRNA.Figure 6. AKR1B1 overexpression reversed the effects of NORAD silencing on proliferation, invasion, and migration of glioma cells. (A) The expression of AKR1B1 was evaluated using RT-qPCR after transfection with AKR1B1 pcDNA3.1. Three independent experiments were carried out (n=3). ** P<0.01 versus pcDNA-NC. (B) CCK-8 assay and (C) colony formation assay tested the proliferation of U251 cells. (D) Invasion of U251 cells was evaluated using Transwell assay (magnification 200×). (E) Representative images of cell migration (magnification 100×). (F) Quantification of cell invasion. (G) Relative quantification, as examined using wound healing assay. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05, ## P<0.01, ### P<0.001 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; pcDNA-AKR1B1 – AKR1B1 overexpressed plasmid pcDNA3.1; pcDNA-NC – empty vector plasmid; CCK-8 – Cell Counting Kit-8; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD.Figure 7. NORAD silencing decreased the expression of migration-related genes in glioma cells. The (A) proteins and (B) mRNA expression of MMP2, MMP9, and vimentin was detected using western blotting and RT-qPCR, respectively. All experiments were repeated three times independently (n=3). ** P<0.01, *** P<0.001 versus si-NC; # P<0.05, ## P<0.01 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; mRNA – messenger RNA; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid.Figure 8. (A–C) AKR1B1 overexpression attenuated the effects of NORAD silencing on apoptosis of glioma cells. (A) Flow cytometry analysis detected the apoptotic rate of U251 cells. (B) The expression of apoptosis-related proteins was examined using western blotting. Three independent experiments were carried out (n=3). ** P<0.01, *** P<0.001 vs. si-NC; # P<0.05, ### P<0.001 versus si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; si-NC – negative control for NORAD siRNA; si-NORAD – si-NORAD, small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid.Figure 9. (A, B) NORAD could regulate ERK signaling via targeting AKR1B1. The expression of p-ERK was measured using western blotting. All experiments were repeated 3 times independently (n=3). *** P<0.001 vs. si-NC; # P<0.05 vs. si-NORAD-2+pcDNA-NC. NORAD – noncoding RNA activated by DNA damage; ERK – extracellular signal-regulated kinase; p-ERK – phosphorylated ERK; si-NC – negative control for NORAD siRNA; si-NORAD – small interfering RNAs (siRNAs) against NORAD; pcDNA-NC – empty vector plasmid. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387