05 October 2020: Animal Study
The Protein Kinase R Inhibitor C16 Alleviates Sepsis-Induced Acute Kidney Injury Through Modulation of the NF-κB and NLR Family Pyrin Domain-Containing 3 (NLPR3) Pyroptosis Signal Pathways
Jialu Zhou1B, Fan Zhang2B, Hongru Lin1B, Minxue Quan1B, Yaqin Yang1B, Yanni Lv3C, Zongnan He4CE, Yisong Qian1ACE*DOI: 10.12659/MSM.926254
Med Sci Monit 2020; 26:e926254






Abstract
BACKGROUND: Protein kinase R (PKR) is implicated in the inflammatory response to bacterial infection while the role of PKR in sepsis-induced acute kidney injury (AKI) is largely unknown. This study aimed to investigate the effects of the specific PKR inhibitor C16 (C13H8N4OS) on lipopolysaccharide (LPS)-induced AKI, and its mechanisms of action.
MATERIAL AND METHODS: C57BL/6J mice were injected intraperitoneally with C16 or vehicle 1 h before the LPS challenge and then injected intraperitoneally with LPS or 0.9% saline. After the LPS challenge, histopathological damage, renal function, and levels of proinflammatory cytokines were assessed. All the related signaling pathways were analyzed.
RESULTS: C16 effectively inhibited LPS-induced renal elevation of proinflammatory cytokines and chemokines. C16 prevented NF-κB activation and suppressed the PKR/eIF2α signaling pathway in AKI after the LPS challenge. Furthermore, C16 significantly inhibited pyroptosis during AKI, as evidenced by decreased renal levels of apoptosis-associated speck-like protein; NACHT, LRR, NLR Family Pyrin Domain-Containing 3; caspase-1; interleukin (IL)-1β; and IL-18.
CONCLUSIONS: Our findings suggest that inhibition by C16 ameliorated LPS-induced renal inflammation and injury, at least partly through modulation of the pyroptosis signal pathway in the kidney.
Keywords: Acute Kidney Injury, Inflammasomes, Lipopolysaccharides, Indoles, NLR Family, Pyrin Domain-Containing 3 Protein, Pyroptosis, Sepsis, Thiazoles, eIF-2 Kinase
Background
Acute kidney injury (AKI) is defined as a sudden and reversible renal dysfunction associated with systemic inflammation [1,2]. Sepsis, induced by a pathogen or an endotoxin, is implicated in the pathogenesis of AKI [3]. Lipopolysaccharide (LPS), a common source for simulating clinical AKI, acts on the toll-like receptor (
Protein kinase R (PKR), originally found to play a role in innate antiviral immune response, senses various stimuli, such as viral double-stranded RNAs, bacterial infection, and inflammatory cytokines [6–8]. The activated PKR leads to phosphorylation of eukaryotic translation initiation factor 2α (eIF2α), thereby suppressing translation initiation as an antiviral strategy [9]. In addition, activated PKR provokes diverse downstream events, such as the activity of inflammasomes and transcription factors in induction of apoptosis and inflammation [10,11]. Recently, evidence has been presented indicating that PKR mediates acute lung injury, whereas pharmacological inhibition of PKR may ameliorate organ damage through anti-inflammatory mechanisms [12,13]. However, the precise role and regulatory mechanism of PKR in the pathogenesis of AKI remain to be elucidated.
Here, we investigated whether inhibition of PKR by the specific inhibitor C16 (C13H8N4OS) reduces acute renal inflammation and injury elicited by the LPS challenge. The possible signaling pathways through which PKR may mediate AKI also were explored.
Material and Methods
REAGENTS:
The PKR inhibitor C16 and lipopolysaccharide (LPS) (
ANIMALS AND LIPOPOLYSACCHARIDE CHALLENGE:
Mice with a C57BL/6J genetic background were provided by the Model Animal Research Center of Yangzhou University (Yangzhou certificate no. SCXK2012-0004). Experimental protocols were approved by the Institutional Animal Care and Use Committee of Nanchang University. All procedures were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Male mice (age ~8 weeks, weight 18 to 22 g) were used in the control and the experimental groups. One hour before the LPS challenge, C16 (0.5 and 1 mg/kg in 0.5% dimethyl sulfoxide [DMSO]) or vehicle (0.5% DMSO in phosphate-buffered saline) was administered intraperitoneally to the animals. Then the mice were subjected to intraperitoneal injection of LPS (15 mg/kg) or an equal volume of 0.9% saline as a control.
HISTOLOGIC ANALYSIS:
The mice were euthanized 24 h after the LPS challenge for histopathological examination. The formalin-fixed kidneys were sliced and stained with hematoxylin and eosin (H&E). Discontinuous regions of the renal cortex were observed and photomicrographs were made using a light microscope. The histological tissue damage was semiquantitatively evaluated and scored using the following scale: no damage, 0; <25% damage, 1; 25% to 50% damage, 2; 50% to 75%, 3; and >75% damage, 4 [14].
MEASUREMENT OF CREATININE AND BLOOD UREA NITROGEN:
Serum was collected from each mouse 24 h after the LPS challenge. The concentrations of creatinine and blood urea nitrogen in the serum were measured using a Roche Cobas C311 Chemistry Analyzer (Roche Diagnostics, Indianapolis, IN, USA). The manufacturer’s instructions for the protocol were followed.
REAL-TIME POLYMERASE CHAIN REACTION:
The mRNA levels of tumor necrosis factor (TNF) α, interleukin (IL)-1β, IL-6, monocyte chemoattractant protein-1 (MCP-1), inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2) were measured in renal tissues with real-time polymerase chain reaction (PCR) as previously described [15]. Samples totaling 50 mg were taken from the same area in the renal cortex regions 6 h after the LPS challenge. The tissues were quick-frozen in liquid nitrogen and RNA was isolated with TRIzol™ Reagent (Life Technologies, Carlsbad, CA, USA). The first-strand cDNAs were synthesized with a One Step PrimeScript™ reverse transcription-PCR Kit (Takara, Liaoning, China). The reaction mixture contained 2× SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA), primer pairs, and cDNA. The reaction consisted of a 2-step thermocycling protocol (40 cycles at 95°C for 15 s and 60°C for 1 min). The mRNA levels were calculated by using the 2−ΔΔCT method. The primer sequences used in the experiment are listed in Table 1.
WESTERN BLOT:
Protein samples were collected from kidneys 6 h after the LPS challenge, and protein content was quantified using a BCA Protein Assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Western blotting was performed as previously described [16]. Briefly, equal amounts of protein were loaded into the wells of the sodium dodecyl sulfate-polyacrylamide electrophoresis gel and separated by electrophoresis for 2 h at 100 V. Then the protein was transferred from the gel to the membrane. Five percent (w/v) dried skim milk powder was used to block nonspecific binding. Membranes were incubated with different primary antibodies at 4°C overnight, followed by incubation for 1 h at room temperature with horseradish peroxidase-conjugated secondary antibodies. The bands were detected with a SuperSignal West Pico Chemiluminescent Substrate Trial Kit (Pierce, Rockford, IL, USA) and analyzed with the ChemiDoc XRS system using Quantity One software (Bio-Rad, Richmond, CA, USA).
ENZYME-LINKED IMMUNOSORBENT ASSAY:
The protein levels of IL-1β and IL-18 were detected in the renal homogenate 6 h after the LPS challenge, using commercial enzyme-linked immunosorbent assay kits (NeoBioscience, Shenzhen, China). The optical density was detected at 450 nm and the protein concentration was obtained from the absorbance curves generated following standards for recombinant proteins.
STATISTICAL ANALYSIS:
Data were obtained from at least 3 independent experiments and were represented as mean±SD. Statistical differences were compared using one-way analysis of variance (ANOVA) followed by a Tukey’s test. An unpaired
Results
C16 AMELIORATES SEPSIS-INDUCED RENAL INJURY AND IMPROVES RENAL FUNCTION IN MICE:
To investigate the protective effects of C16, H&E staining was used to detect histological changes in renal tissues. In the vehicle-treated group, the renal tissues exhibited normal morphology. Exposure to LPS resulted in obvious tubular swelling, tubular dilatation and distortion, necrosis (with condensed or missing nuclei), interstitial hemorrhage, and infiltration of inflammatory cells. However, pretreatment with C16 (0.5 and 1 mg/kg) significantly reduced histopathological alterations in the LPS-challenged mice (Figure 1A). Quantitative analysis further revealed that renal damage scores were significantly improved in C16-treated mice compared with control mice after the LPS challenge (Figure 1B). In addition, the mice treated with C16 also exhibited significantly lower levels of creatinine and blood urea nitrogen compared with the LPS-treated mice (Figure 1C, 1D). These data suggest that C16 is effective in attenuating AKI and improving renal function after LPS stimulation.
C16 INHIBITS RENAL INFLAMMATORY RESPONSES INDUCED BY LPS:
To confirm whether C16 regulates renal inflammatory responses, we performed real-time PCR to detect mRNA levels of proinflammatory genes following LPS stimulation. As expected, remarkable elevations in TNFα, IL-1β, IL-6, MCP-1, iNOS, and COX-2 were observed 6 h after LPS injection. In contrast, both doses of C16 treatment suppressed renal expression of these cytokines (Figure 2).
C16 PREVENTS LPS-INDUCED NF-κB ACTIVATION IN AKI:
Because NF-κB is one of the most important signal pathways that mediate cellular inflammatory response, we used western blotting to assess whether C16 has effects on NF-κB activation in AKI. LPS induced phosphorylation of IKKα/β, p65, and IκBα and caused IκBα degradation, indicating that the NF-κB signal pathway had been activated (Figure 3A). C16 significantly decreased levels of phospho-IKKα/β (Figure 3A, 3B), phospho-p65 (pp65) (Figure 3A, 3C) and phospho-IκBα, Figure 3A, 3D), and suppressed degradation of IκBα (Figure 3A, 3E) after LPS stimulation. These results suggest that C16 is a potent inhibitor of NF-κB activation in AKI.
C16 SUPPRESSES THE PKR/EIF2α SIGNALING PATHWAY IN THE KIDNEY:
The PKR/eIF2α signaling pathway is essential in infection and innate immunity. Phosphorylation of eIF2α by PKR induces attenuation of translation and reduces inhibition of NF-κB, which in turn promotes cytokine production [17]. We investigated whether the anti-inflammatory effects of the PKR inhibitor were associated with modulation of the PKR/eIF2α signaling pathway. LPS promoted phosphorylation of PKR T451 residue whereas this effect was suppressed by treatment with C16 (Figure 4A, 4B). The PKR-activated phosphorylation of eIF2α also was reversed in the presence of C16 (Figure 4A, 4C).
C16 INHIBITS LPS-INDUCED PYROPTOSIS AFTER LPS CHALLENGE:
Pyroptosis is a recently described type of inflammatory programmed cell death. Pyroptosis is often triggered by infection or various pathological stimuli, and eventually exacerbates inflammation [18,19]. Activation of the NACHT, LRR, and Pyrin domains-containing protein 3 (NLRP3) inflammasome is involved with AKI and the process of pyroptosis [20]. Emerging evidence indicates that PKR mediates NLRP3 activation [21]. Therefore, we investigated the effect of C16 on the NLRP3-related pyroptosis pathway. Western blot showed that LPS induced expression of NLRP3, the ASC (apoptosis-associated speck-like protein containing a caspase recruitment domain), and cleaved caspase-1, which were all suppressed by treatment with C16 (Figure 5A–5D). Treatment with C16 also decreased LPS-induced IL-1β and IL-18 levels in the kidneys (Figure 5E, 5F).
Discussion
We demonstrated that the PKR inhibitor C16 significantly attenuates sepsis-induced AKI, improves renal function, and ameliorates renal inflammatory response in a mouse model of LPS injection. Mechanistically, C16 prevents NF-κB activation, inhibits the PKR/eIF2α signal pathway, and regulates NLRP3-related pyroptosis in the kidney, which suggests that PKR may be a potential target in renal inflammation and injury.
PKR was previously implicated in the defense mechanism against viruses, and recently has been found to be a major regulator of transcriptional control, mRNA translation, cell proliferation, and apoptosis [22]. PKR can be activated by various endogenous and exogenous factors, including pathogens, endotoxins, cytokines, irradiation, and multiple stresses [23,24]. Inhibition of PKR has been proven to have beneficial effects in LPS-induced organ injury and systematic inflammation. PKR inhibition has been shown to decrease inflammation and attenuate lung injury in LPS-challenged mice [12,13]. The present
Our results suggested that the protective effect of C16 on AKI is closely related to renal inflammation. Therefore, we explored the signal pathway-mediating inflammatory response. PKR was shown to activate the proinflammatory NF-κB pathway by directly interacting with the IKK complex in the presence of small amounts of double-strand RNA in cells, such as at the onset of viral infection [27]. By using the PKR inhibitor C16, we confirmed that inhibition of PKR also prevents NF-κB activation in LPS-challenged renal tissue. Thus, we hypothesize that C16 prevents NF-κB activation and suppresses transcription of NF-κB target genes, which in turn reduces inflammatory response during AKI.
PKR is one of the kinases that regulates phosphorylation of eukaryotic initiation factor 2 on its α subunit (eIF2α), a major regulator of the initiation phase of mRNA translation. Phosphorylation of eIF2α by PKR leads to the transient suppression of general protein synthesis, resulting in a decrease in or prevention of viral replication [22]. The inhibitory effects of C16 on the PKR/eIF2α signal pathway were confirmed in our study. However, how eIF2α modulates LPS-induced AKI and inflammation, possibly through mediation of endoplasmic reticulum stress [28], remains to be elucidated.
Pyroptosis is a recently identified form of programmed cell death and NLRP3 is a key factor in this process [29]. NLRP3 activation is usually triggered by pathogen-associated molecular patterns (PAMPs) or endogenous infection in the host cells, and promotes pro-caspase-1 recruitment [30]. Activated caspase-1 (the cleaved form) proteolytically cleaves the cytokine precursors pro-IL-1β and pro-IL-18, which are critical for the release of the biologically active forms (IL-1β and IL-18), and consequently triggers a proinflammatory response [31]. PKR was previously believed to regulate the activity of the NLRP3 inflammasome [10]. PKR inhibition was shown to prevent PKR activation and inhibit NLRP3-mediated pyroptosis in LPS-challenged macrophages [12]. In the present study, we investigated whether PKR inhibition influences the NLRP3 pyroptosis pathway in AKI. In the
Conclusions
In summary, the PKR inhibitor C16 acts on the PKR/eIF2α signal pathway to inhibit NF-κB activation and suppresses the NLRP3 pyroptosis signal pathway, which are both associated with mediation of inflammatory responses. The result is attenuation of LPS-induced renal inflammation and injury. Pharmacological inhibition of PKR may be a novel therapeutic strategy for sepsis-induced AKI.
Figures
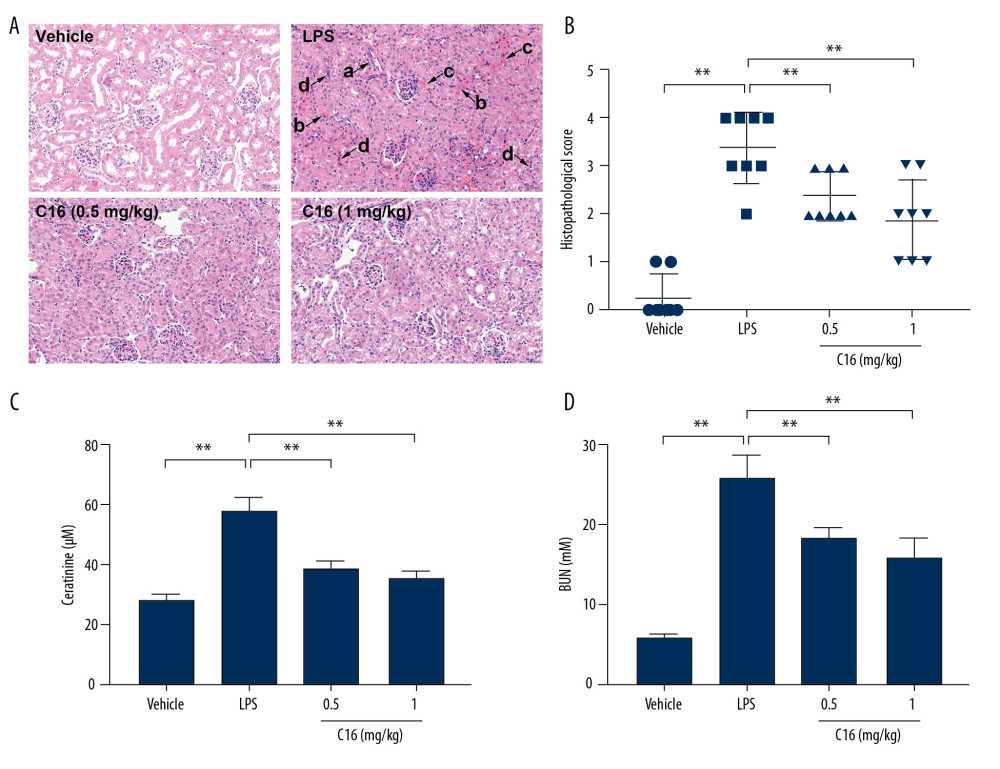 Figure 1. C16 ameliorates sepsis-induced acute kidney injury in mice. (A) Representative hematoxylin and eosin-stained paraffin sections of kidney. Damage is indicated by arrows. (a: Tubular dilation and distortion. b: Necrosis. c: Interstitial hemorrhage. d: Infiltration of inflammatory cells.) Scale bar, 50 μm. (B) Histopathological score for the kidney. (C) Blood urea nitrogen in serum. (D) Creatinine levels in serum. Data are shown as mean ± standard deviation (n=8). ** P<0.01.
Figure 1. C16 ameliorates sepsis-induced acute kidney injury in mice. (A) Representative hematoxylin and eosin-stained paraffin sections of kidney. Damage is indicated by arrows. (a: Tubular dilation and distortion. b: Necrosis. c: Interstitial hemorrhage. d: Infiltration of inflammatory cells.) Scale bar, 50 μm. (B) Histopathological score for the kidney. (C) Blood urea nitrogen in serum. (D) Creatinine levels in serum. Data are shown as mean ± standard deviation (n=8). ** P<0.01. 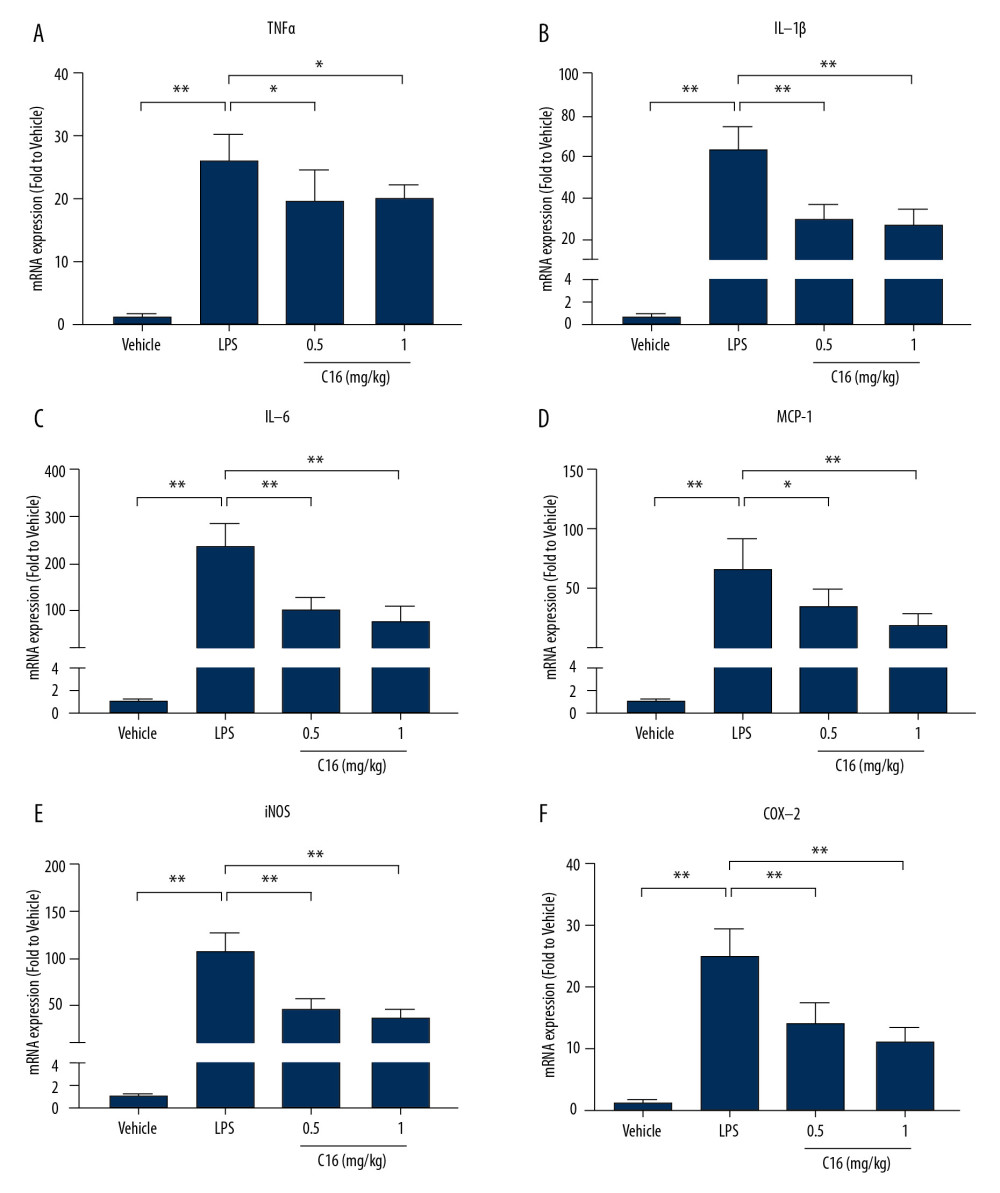 Figure 2. C16 inhibits renal inflammatory responses after lipopolysaccharide (LPS) injection. Real-time polymerase chain reaction was used to measure renal levels of proinflammatory cytokines and chemokines after LPS injection. (A) Tumor necrosis factor α. (B) Interleukin (IL)-1β. (C) IL-6. (D) Monocyte chemoattractant protein-1. (E) inducible nitric oxide synthase. (F) Cyclooxygenase-2. Data are presented as mean±standard deviation (n=6). * P<0.05, ** P<0.01.
Figure 2. C16 inhibits renal inflammatory responses after lipopolysaccharide (LPS) injection. Real-time polymerase chain reaction was used to measure renal levels of proinflammatory cytokines and chemokines after LPS injection. (A) Tumor necrosis factor α. (B) Interleukin (IL)-1β. (C) IL-6. (D) Monocyte chemoattractant protein-1. (E) inducible nitric oxide synthase. (F) Cyclooxygenase-2. Data are presented as mean±standard deviation (n=6). * P<0.05, ** P<0.01. 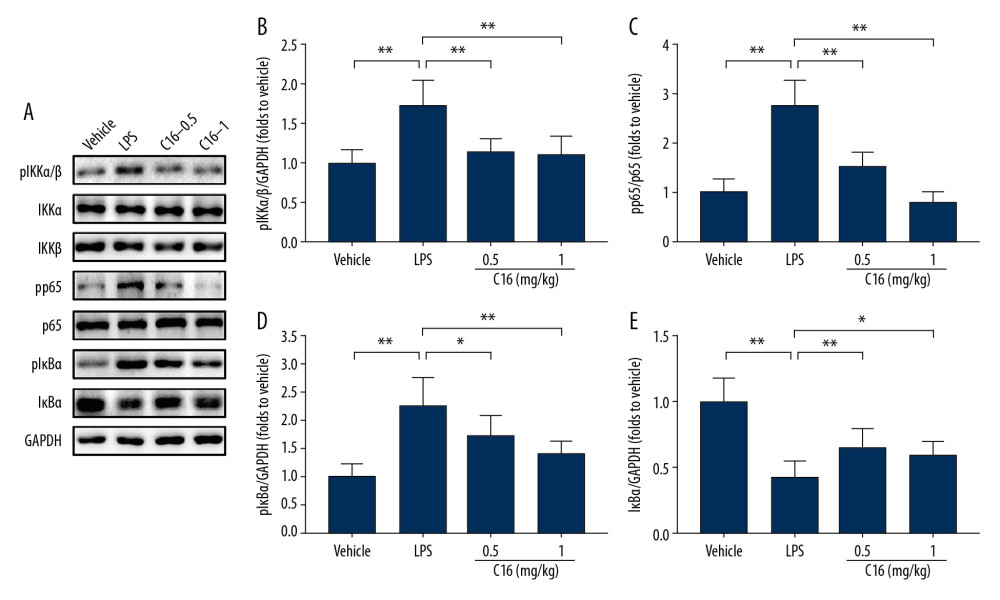 Figure 3. C16 prevents NF-κB activation in lipopolysaccharide-induced acute kidney injury. (A) Immunoblots of phospho-IKKα/β (pIKKα/β), total IKKα (IKKα), total IKKβ (IKKβ), phospho-NF-κB p65 (pp65), total NF-κB p65 (p65), phospho-IκBα (pIκBα), total IκBα (IκBα), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pIKKα/β, (C) pp65, (D) pIκBα, and (E) IκBα were quantitatively analyzed and are shown as mean± standard deviation (n=6). * P<0.05, ** P<0.01.
Figure 3. C16 prevents NF-κB activation in lipopolysaccharide-induced acute kidney injury. (A) Immunoblots of phospho-IKKα/β (pIKKα/β), total IKKα (IKKα), total IKKβ (IKKβ), phospho-NF-κB p65 (pp65), total NF-κB p65 (p65), phospho-IκBα (pIκBα), total IκBα (IκBα), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pIKKα/β, (C) pp65, (D) pIκBα, and (E) IκBα were quantitatively analyzed and are shown as mean± standard deviation (n=6). * P<0.05, ** P<0.01. 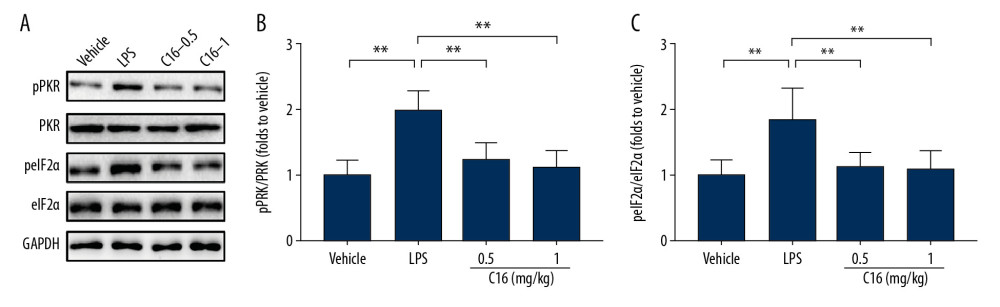 Figure 4. C16 suppresses the renal protein kinase R (PKR)/eIF2α signaling pathway. (A) Immunoblots of phospho-PKR (pPKR), total PKR (PKR), phospho-eIF2α (peIF2α), total eIF2α (eIF2α), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pPKR and (C) peIF2α were quantitatively analyzed and are shown as mean±standard deviation (n=8). **P<0.01.
Figure 4. C16 suppresses the renal protein kinase R (PKR)/eIF2α signaling pathway. (A) Immunoblots of phospho-PKR (pPKR), total PKR (PKR), phospho-eIF2α (peIF2α), total eIF2α (eIF2α), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pPKR and (C) peIF2α were quantitatively analyzed and are shown as mean±standard deviation (n=8). **P<0.01. 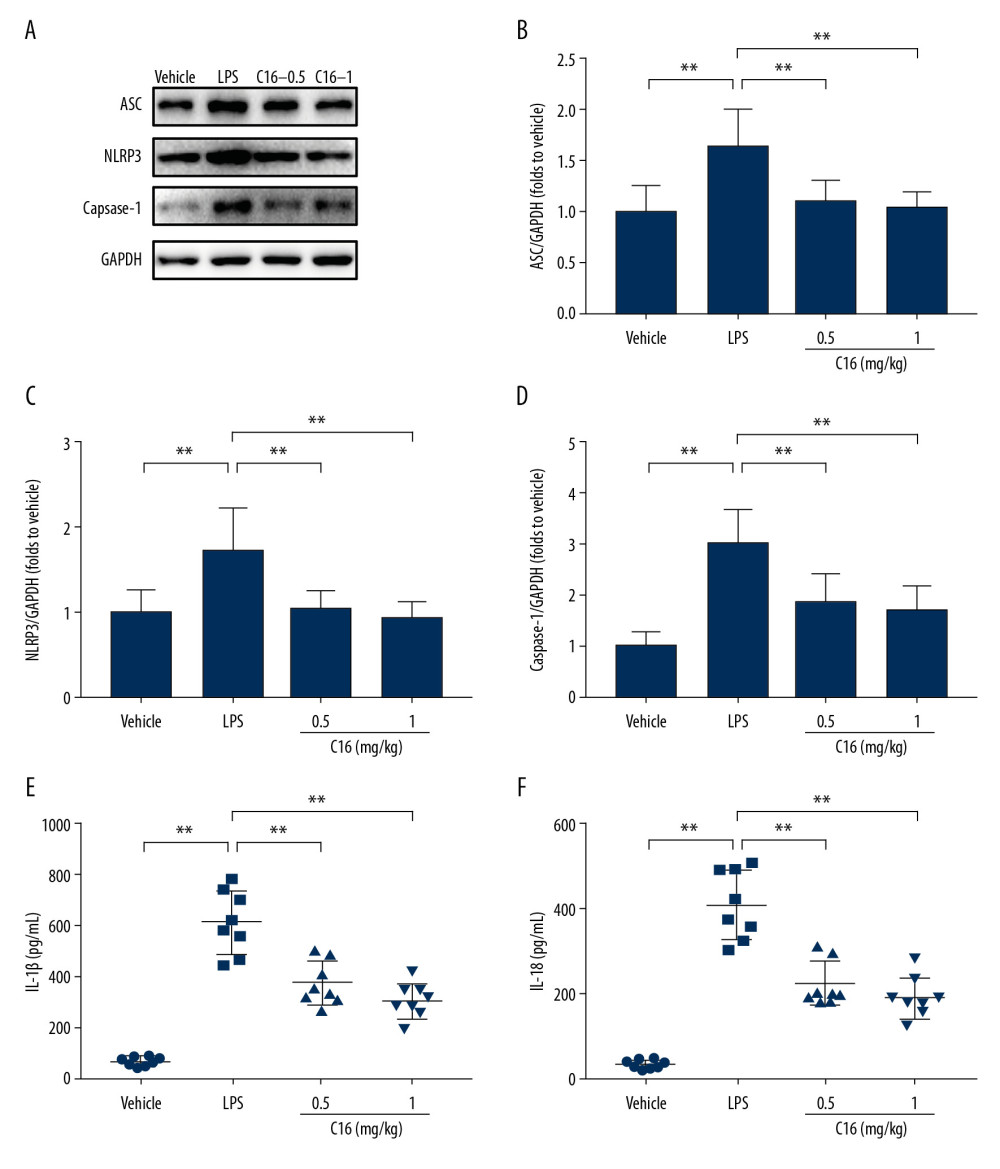 Figure 5. C16 inhibits lipopolysaccharide (LPS)-induced pyroptosis after LPS challenge. (A) Immunoblots of apoptosis-associated speck-like protein (ASC); NACHT, LRR, NLR Family Pyrin Domain-Containing 3 (NLRP3); caspase-1 (cleaved form); and glyceraldehyde-3-phosphate dehydrogenase. We quantitatively analyzed levels of (B) ASC, (C) NLRP3, and (D) cleaved caspase-1. (E) The production of interleukin (IL)-1β in the kidney was detected with enzyme-linked immunosorbent assay (ELISA). (F) The production of IL-18 in the kidney was detected with ELISA. All values are shown as mean±standard deviation (n=8). ** P<0.01.
Figure 5. C16 inhibits lipopolysaccharide (LPS)-induced pyroptosis after LPS challenge. (A) Immunoblots of apoptosis-associated speck-like protein (ASC); NACHT, LRR, NLR Family Pyrin Domain-Containing 3 (NLRP3); caspase-1 (cleaved form); and glyceraldehyde-3-phosphate dehydrogenase. We quantitatively analyzed levels of (B) ASC, (C) NLRP3, and (D) cleaved caspase-1. (E) The production of interleukin (IL)-1β in the kidney was detected with enzyme-linked immunosorbent assay (ELISA). (F) The production of IL-18 in the kidney was detected with ELISA. All values are shown as mean±standard deviation (n=8). ** P<0.01. 
References
1. Hoste EAJ, Kellum JA, Selby NM, Global epidemiology and outcomes of acute kidney injury: Nat Rev Nephrol, 2018; 14(10); 607-25
2. Uchino S, Kellum JA, Bellomo R, Acute renal failure in critically ill patients: A multinational, multicenter study: JAMA, 2005; 294(7); 813-18
3. Alobaidi R, Basu RK, Goldstein SL, Bagshaw SM, Sepsis-associated acute kidney injury: Semin Nephrol, 2015; 35(1); 2-11
4. Niu X, Yao Q, Li W, Harmine mitigates LPS-induced acute kidney injury through inhibition of the TLR4-NF-kappaB/NLRP3 inflammasome signalling pathway in mice: Eur J Pharmacol, 2019; 849; 160-69
5. Chen Y, Jin S, Teng X, Hydrogen sulfide attenuates LPS-induced acute kidney injury by inhibiting inflammation and oxidative stress: Oxid Med Cell Longev, 2018; 2018 6717212
6. Meurs E, Chong K, Galabru J, Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon: Cell, 1990; 62(2); 379-90
7. Lee YS, Kunkeaw N, Lee YS, Protein kinase R and its cellular regulators in cancer: An active player or a surveillant? WIREs RNA: Wiley Interdiscip Rev RNA, 2020; 11(2); e1558
8. Xu H, Chen J, Si X, PKR inhibition mediates endotoxin tolerance in macrophages through inactivation of PI3K/AKT signaling: Mol Med Rep, 2018; 17(6); 8548-56
9. Ilan L, Osman F, Namer LS, PKR activation and eIF2alpha phosphorylation mediate human globin mRNA splicing at spliceosome assembly: Cell Res, 2017; 27(5); 688-704
10. Lu B, Nakamura T, Inouye K, Novel role of PKR in inflammasome activation and HMGB1 release: Nature, 2012; 488(7413); 670-74
11. Zhang P, Samuel CE, Induction of protein kinase PKR-dependent activation of interferon regulatory factor 3 by vaccinia virus occurs through adapter IPS-1 signaling: J Biol Chem, 2008; 283(50); 34580-87
12. Zeng Y, Qin Q, Li K, PKR suppress NLRP3-pyroptosis pathway in lipopolysaccharide-induced acute lung injury model of mice: Biochem Biophys Res Commun, 2019; 519(1); 8-14
13. Li Y, Xiao J, Tan Y, Inhibition of PKR ameliorates lipopolysaccharide-induced acute lung injury by suppressing NF-kappaB pathway in mice: Immunopharmacol Immunotoxicol, 2017; 39(4); 165-72
14. Liu T, Fang Y, Liu S, Limb ischemic preconditioning protects against contrast-induced acute kidney injury in rats via phosphorylation of GSK-3beta: Free Radic Biol Med, 2015; 81; 170-82
15. Guan T, Qian Y, Tang X, Maslinic acid, a natural inhibitor of glycogen phosphorylase, reduces cerebral ischemic injury in hyperglycemic rats by GLT-1 up-regulation: J Neurosci Res, 2011; 89(11); 1829-39
16. Huang L, Guan T, Qian Y, Anti-inflammatory effects of maslinic acid, a natural triterpene, in cultured cortical astrocytes via suppression of nuclear factor-kappa B: Eur J Pharmacol, 2011; 672(1–3); 169-74
17. Fung TS, Liu DX, Coronavirus infection, ER stress, apoptosis and innate immunity: Front Microbiol, 2014; 5; 296
18. Man SM, Karki R, Kanneganti TD, Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases: Immunol Rev, 2017; 277; 61-75
19. Fenini G, Karakaya T, Hennig P, Pyroptosis: Host cell death and inflammation: Nat Rev Microbiol, 2009; 7(2); 99-109
20. Anders HJ, Muruve DA, The inflammasomes in kidney disease: J Am Soc Nephrol, 2011; 22(6); 1007-18
21. Li Y, Li H, Liu S, Pirfenidone ameliorates lipopolysaccharide-induced pulmonary inflammation and fibrosis by blocking NLRP3 inflammasome activation: Mol Immunol, 2018; 99; 134-44
22. Gal-Ben-Ari S, Barrera I, Ehrlich M, Rosenblum K, PKR: A kinase to remember: Front Mol Neurosci, 2019; 11; 480
23. Garcia MA, Meurs EF, Esteban M, The dsRNA protein kinase PKR: Virus and cell control: Biochimie, 2007; 89(6–7); 799-811
24. Watanabe T, Imamura T, Hiasa Y, Roles of protein kinase R in cancer: Potential as a therapeutic target: Cancer Sci, 2018; 109(4); 919-25
25. Jiang Y, Steinle JJ, Epac1 inhibits PKR to reduce NLRP3 inflammasome proteins in retinal endothelial cells: J Inflamm Res, 2019; 12; 153-59
26. Jammi NV, Whitby LR, Beal PA, Small molecule inhibitors of the RNA-dependent protein kinase: Biochem Biophysl Res Commun, 2003; 308(1); 50-57
27. Bonnet MC, Weil R, Dam E, PKR stimulates NF-kappaB irrespective of its kinase function by interacting with the IkappaB kinase complex: Mol Cellular Biol, 2000; 20(13); 4532-42
28. Palomera-Avalos V, Grinan-Ferre C, Izquierdo V, Resveratrol modulates response against acute inflammatory stimuli in aged mouse brain: Exp Gerontol, 2018; 102; 3-11
29. Broz P, Dixit VM, Inflammasomes: Mechanism of assembly, regulation and signalling: Nat Rev Immunol, 2016; 16(7); 407-20
30. Aachoui Y, Sagulenko V, Miao EA, Stacey KJ, Inflammasome-mediated pyroptotic and apoptotic cell death, and defense against infection: Curr Opin Microbiol, 2013; 16(3); 319-26
31. Martinon F, Tschopp J, Inflammatory caspases: Linking an intracellular innate immune system to autoinflammatory diseases: Cell, 2004; 117(5); 561-74
Figures
Figure 1. C16 ameliorates sepsis-induced acute kidney injury in mice. (A) Representative hematoxylin and eosin-stained paraffin sections of kidney. Damage is indicated by arrows. (a: Tubular dilation and distortion. b: Necrosis. c: Interstitial hemorrhage. d: Infiltration of inflammatory cells.) Scale bar, 50 μm. (B) Histopathological score for the kidney. (C) Blood urea nitrogen in serum. (D) Creatinine levels in serum. Data are shown as mean ± standard deviation (n=8). ** P<0.01.Figure 2. C16 inhibits renal inflammatory responses after lipopolysaccharide (LPS) injection. Real-time polymerase chain reaction was used to measure renal levels of proinflammatory cytokines and chemokines after LPS injection. (A) Tumor necrosis factor α. (B) Interleukin (IL)-1β. (C) IL-6. (D) Monocyte chemoattractant protein-1. (E) inducible nitric oxide synthase. (F) Cyclooxygenase-2. Data are presented as mean±standard deviation (n=6). * P<0.05, ** P<0.01.Figure 3. C16 prevents NF-κB activation in lipopolysaccharide-induced acute kidney injury. (A) Immunoblots of phospho-IKKα/β (pIKKα/β), total IKKα (IKKα), total IKKβ (IKKβ), phospho-NF-κB p65 (pp65), total NF-κB p65 (p65), phospho-IκBα (pIκBα), total IκBα (IκBα), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pIKKα/β, (C) pp65, (D) pIκBα, and (E) IκBα were quantitatively analyzed and are shown as mean± standard deviation (n=6). * P<0.05, ** P<0.01.Figure 4. C16 suppresses the renal protein kinase R (PKR)/eIF2α signaling pathway. (A) Immunoblots of phospho-PKR (pPKR), total PKR (PKR), phospho-eIF2α (peIF2α), total eIF2α (eIF2α), and glyceraldehyde-3-phosphate dehydrogenase. The levels of (B) pPKR and (C) peIF2α were quantitatively analyzed and are shown as mean±standard deviation (n=8). **P<0.01.Figure 5. C16 inhibits lipopolysaccharide (LPS)-induced pyroptosis after LPS challenge. (A) Immunoblots of apoptosis-associated speck-like protein (ASC); NACHT, LRR, NLR Family Pyrin Domain-Containing 3 (NLRP3); caspase-1 (cleaved form); and glyceraldehyde-3-phosphate dehydrogenase. We quantitatively analyzed levels of (B) ASC, (C) NLRP3, and (D) cleaved caspase-1. (E) The production of interleukin (IL)-1β in the kidney was detected with enzyme-linked immunosorbent assay (ELISA). (F) The production of IL-18 in the kidney was detected with ELISA. All values are shown as mean±standard deviation (n=8). ** P<0.01. In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387