18 October 2020: Animal Study
Silencing of Long Non-Coding RNA X Inactive Specific Transcript (Xist) Contributes to Suppression of Bronchopulmonary Dysplasia Induced by Hyperoxia in Newborn Mice via microRNA-101-3p and the transforming growth factor-beta 1 (TGF-β1)/Smad3 Axis
Wenhao Yuan1ABCDEF, Xiaoyan Liu1ABCDG, Lingkong Zeng1BCDFG*, Hanchu Liu1ABCDEFG, Baohuan Cai2ABCDEFG, Yanping Huang1ABDF, Xuwei Tao1ABCDF, Luxia Mo1ABDF, Lingxia Zhao1BCDF, Chunfang Gao1BCDFDOI: 10.12659/MSM.922424
Med Sci Monit 2020; 26:e922424






Abstract
BACKGROUND: Bronchopulmonary dysplasia (BPD) is a chronic lung disease mostly affecting premature infants. Long non-coding RNA (lncRNA) X inactive specific transcript (Xist) is actively involved in pulmonary disease development. The present study explored the potential mechanism of Xist in BPD development.
MATERIAL AND METHODS: First, newborn BPD mouse models were successfully established. lncRNAs and genes with differential expression were identified using microarray analysis. Various injuries and radial alveolar counts of lung tissues of BPD mice were detected by hematoxylin-eosin staining. Functional assays were utilized to detect alterations of superoxide dismutase (SOD), malondialdehyde (MDA), vascular endothelial growth factor, collagen I, alpha-smooth muscle Actin, TGF-β1, and Smad3. Then, dual-luciferase reporter gene assay and RNA pull-down assay were performed to clarify the targeting relationship between Xist and miR-101-3p and between miR-101-3p and high-mobility group protein B3 (HMGB3).
RESULTS: In BPD mice, radial alveolar counts value and SOD activity declined while MDA level increased. Results of microarray analysis found that Xist and HMGB3 were highly expressed in BPD mice. Next, silenced Xist alleviated lung damage in BPD mice. Xist competitively bound to miR-101-3p to activate HMGB3, and overexpressed miR-101-3p mitigated lung damage in BPD mice. Additionally, silenced Xist downregulated the TGF-β1/Smad3 axis.
CONCLUSIONS: Our study demonstrated that silencing of Xist suppressed BPD development by binding to miR-101-3p and downregulating HMGB3 and the TGF-b1/Smad3 axis. Our results may provide novel insights for BPD treatment.
Keywords: Bronchopulmonary Dysplasia, HMGB3 Protein, Animals, Newborn, Gene Silencing, Hyperoxia, Smad3 Protein
Background
Bronchopulmonary dysplasia (BPD) is a severe and common chronic respiratory disease among premature infants, which causes potentially life-long effects on normal respiratory functions [1]. BPD mostly afflicts premature infants and has dangerously high incidence and mortality rates [2]. The mechanism of BPD pathogenesis is complex and diverse, including hereditary factors, hyperoxia, ante- and postnatal sepsis, and invasive mechanical ventilation [3]. Hyperoxia-mediated oxidative stress can enhance critical lung stem cell apoptosis and suppress its growth, promoting BPD progression [4]. Pulmonary fibrosis and alveolar septation suspension are also related to BPD [5]. In addition to damaging lung functions of premature infants, BPD also substantially impedes the normal growth of lung functions, even in later childhood, and causes high susceptibility to asthma [6]. Therefore, biomarkers in the early stage and novel therapeutic strategies for BPD are urgently needed. The present study assessed the interaction between oxidative stress and lung functions to develop novel intervention strategies.
Although autopsy on human BPD corpses is more informative, the increased survival rates of BPD patients makes it hard to obtain such corpses; therefore, BPD experiments are mostly performed on animal models, such as mice, rats, lambs, and rabbits [7]. Since the lung tissue morphology and gene expression are quite different between rats and newborn mice, newborn mice are more often used in BPD modeling [3]. Hyperoxia is the most common method to establish BPD mouse models [8]. Therefore, mice are considered as the most suitable animals for BPD modeling given their fast growth and wide availability [7].
Long non-coding RNAs (lncRNAs) are important in disease occurrence and progression, and exploration of their associations with diseases contribute insightful perspectives about the pathogenesis, diagnosis, and therapies of diverse human diseases [9]. Abundant evidence suggests that differentially expressed lncRNAs actively participate in BPD. It was found that alterations of many lncRNAs are observed in BPD mouse models, which greatly modulates the gene activities, including inflammatory response and oxidative stress, in pulmonary development [10,11]. However, there has been little research on the interaction between Xist and BPD. X inactive specific transcript (Xist) is one of the first identified lncRNAs related to malignancy of many human cancers, including lung cancer [12]. In non-small cell lung cancer (NSCLC) tissues and cells, Xist is overexpressed, which encourages cancer cell growth and inhibits cell apoptosis, leading to active tumor growth [13]. Furthermore, Xist acts as a competitive endogenous RNA (ceRNA) to encourage NSCLC cell growth and invasion [14]. According to previous research, activated Xist can target miR-140 to enhance cell proliferation and metastasis and reduce lung cancer cell apoptosis, increasing BPD progression [12]. miRs are small RNA molecules which silence genes and inhibit translation through binding to their target RNAs [15]. According to a previous study, many miRs are differentially expressed in BPD, serving as biological targets in BPD research [16]. Based on all the evidence presented above, it is reasonable to hypothesize that Xist also functions as a ceRNA in BPD. Thus, we conducted a series of experiments to test this hypothesis.
Material and Methods
ETHICS STATEMENT:
This study was approved and supervised by the Ethics Committee of Wuhan Children’s Hospital. Significant efforts were made to minimize the number of animals and their suffering.
EXPERIMENTAL ANIMALS AND MODEL PREPARATION:
Ninety-six C57BL/6 healthy newborn mice (3 days old, 3.52±0.25 g) were obtained from Guangzhou University of Chinese Medicine, Guangzhou, Guangdong, China, SYXK (Guangdong, 2019-0202) and were nursed by female mice and raised in standard conditions.
The newborn mice were assigned to a control group (n=16) or a BPD group (n=80). Mice in the BPD group were kept in a custom-made oxygen tank equipped with an inlet/outlet and an oxygen inspection hole, maintained at 24–26°C with constant humidity (50~60%). Fraction of inspiration (Fi) O2 in the tank was kept at 65% and CO2 (absorbed using soda lime) was less than 0.5%. The oxygen tank was opened once a day at 8: 30 a.m. for replenishing food and water and to replace bedding. Newborn mice in the control group were raised in an identical manner with 21% FiO2. The newborn mice in each group were nursed by 2 female mice, and the female mice of each group were exchanged every day. All newborn mice survived the BPD modeling. On the 7th day after BPD modeling, 8 newborn mice were taken from each group and weighed for observation. After that, these mice were euthanized and their lung tissues were detached and used for index measurement. On the 14th day after BPD modeling, 8 newborn mice were taken from each group to repeat these steps.
EXPERIMENTAL GROUPING:
On the 14th day after BPD modeling, the remaining 64 newborn mice were randomly split into 8 groups according to body weight, with 8 mice in each group. On the 14th and the 16th days after BPD modeling, these BPD mice were (1) injected with lentivirus (LV)-short hairpin (sh) negative control (NC) and LV-shXist via the tail vein, (2) intraperitoneally injected with antagomir NC, miR-101-3p antagomir, agomir NC, and miR-101-3p agomir, or (3) were intraperitoneally injected with antagomir NC or miR-101-3p antagomir after an injection of LV-shXist. The specific grouping and number of mice using in each group are listed in Supplementary Figure 1.
Lv-shXist referred to the designed Xist-targeting shRNA sequence inserted into LV pHBLV-U6-Scramble-Luc-Puro vector (Hanbio, Shanghai, China). Antagomir NC, miR-101-3p antagomir, agomir NC, and miR-101-3p agomir were purchased from Guangzhou RiboBio Co. (Guangzhou, Guangdong, China). The newborn mice in each group were nursed by 2 female mice, and the female mice of each group were exchanged every day.
SAMPLE PREPARATION:
On the 7th and the 14th days after BPD modeling, 8 newborn mice from the control group and the BPD group were weighed and then euthanized, after which lung tissues were removed and separated. In each group, 5 pairs of right upper lobes were immediately fixed with 4% paraformaldehyde for 24 h and sliced into paraffin-embedded sections for histological staining. The remaining 3 pairs were used to make tissue homogenate. On the 21st day after BPD modeling, all remaining mice in the BPD group were euthanized. Then, 5 pairs of right upper lobes in each group were routinely made into paraffin-embedded sections for histological staining. The remaining 3 pairs were made into tissue homogenate.
ENZYME-LINKED IMMUNOSORBENT ASSAY (ELISA):
After the tissue homogenate was centrifuged, SOD activity and MDA level in the supernatant were assessed according to the instructions of the SOD, MDA, and ELISA kits (NanJing JianCheng Bioengineering Institute, Nanjing, Jiangsu, China).
LNCRNA MICROARRAY ANALYSIS:
The separated lung tissue samples from the newborn mice were analyzed by Arraystar LncRNAs microarray (Kangcheng Biotech Co., Shanghai, China) and scanned by a GenePix 4000B microarray scanner (Molecular Devices, Sunnyvale, CA, USA). Then, the data were analyzed by Agilent GeneSpring GX (Agilent, CA, USA). According to the standardized data, the fold change between the BPD group and the control group represented the alternation of lncRNAs. Subsequently, the
HEMATOXYLIN-EOSIN (HE) STAINING:
The dewaxed and hydrated paraffin-embedded sections were stained with HE (Beyotime Biotechnology Co., Shanghai, China) and sealed for observation of morphological structure in lung tissues from the newborn mice under an optical microscope (Olympus Optical Co., Tokyo, Japan). The number of alveoli displayed on the perpendicular from the central part of bronchiole to the nearest fibrous septa or pleura, known as radial alveolar counts (RAC), were observed in the stained lung tissue sections under an optical microscope. On the 7th day and the 14th day, 5 sections were taken out from each group, and 5 visual fields in each section were randomly selected to get the average.
REVERSE TRANSCRIPTION-QUANTITATIVE POLYMERASE CHAIN REACTION (RT-QPCR):
TRIzol kits (Invitrogen, Carlsbad, CA, USA) were used to extract the total RNA of lung tissue homogenate, and the extracted RNA was reverse-transcribed into cDNA using SYBR Premix Ex Tap™ II kits (Takara, Kyoto, Japan). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as the internal reference, and the 2−ΔΔCT method was used to calculate relative expression. The primer sequences are shown in Table 1.
WESTERN BLOT ANALYSIS:
The total protein in tissue homogenate of each group was extracted, and its concentration was measured. Proteins were transferred onto the polyvinylidene fluoride membranes after sodium dodecyl sulfate polyacrylamide gel electrophoresis. Next, the membranes were sealed by 5% skim milk powder for 1 h and incubated with the primary antibodies (all from Abcam, Inc., Cambridge, MA, USA), vascular endothelial growth factor (VEGF) (ab32152, 1: 1000), collagen I (ab6308, 1 μg/mL), alpha-smooth muscle Actin (α-SMA) (ab108424, 1: 1000), high-mobility group protein 3 (HMGB3) (ab75782, 1: 2500), TGF-β1 (ab92486, 1 μg/mL), drosophila mothers against decapentaplegic 3 (Smad3) (ab40854, 1: 1000), and p-Smad3 (ab63403, 1: 500) at 4°C overnight. After that, the membranes were cultivated with horseradish peroxidase-labeled goat anti-rabbit immunoglobulin G antibody (ab6728, 1: 2000) at room temperature for 1 h. After exposure, the membranes were photographed using a gel imaging system. GAPDH (ab8245, 1: 500) served as the internal reference, and the ratio of the gray value of the target protein to GAPDH was used as the relative protein expression.
DUAL-LUCIFERASE REPORTER GENE ASSAY:
Binding sites between Xist and miR-101-3p and between miR-101-3p and HMGB3 were certified using online software
RNA PULL-DOWN ASSAY:
After incubation with cytoplasmic extracting solution, miR-101-3p-labeled by biotin was resuspended with lysis buffer on ice for 10 min and centrifuged at 10 000 g for 10 min. Streptavidin beads were sealed with lysis solution containing yeast tRNA and bovine serum albumin at 4°C for 2 h, followed by washing twice in 1 mL lysis solution. Next, streptavidin beads were added with lysis solution and then incubated at 4°C for 4 h, followed by 2 washes in 1 mL lysis solution, 3 washes in low salt buffer, and 1 wash in high salt buffer. TRIzol kits (Invitrogen) were used to separate RNA adhered on beads. RT-qPCR was performed to determine Xist expression.
STATISTICAL ANALYSIS:
SPSS 21.0 (IBM Corp. Armonk, NY, USA) was used for data analysis. Kolmogorov-Smirnov testing indicated whether the data were in normal distribution. Data are shown as mean±standard deviation. The
Results
IN BPD MICE, RAC VALUE AND SOD ACTIVITY DECREASED WHILE MDA LEVEL INCREASED:
On the 7th day and the 14th day, newborn mice in the control group were healthy and rapidly growing, while mice in the BPD group weighed less and were in worse condition (p<0.05) (Figure 1A). Regular alveoli interval was observed in the control group, while expanded alveoli interval and alveolar spaces, as well as confluent alveoli, were found in the BPD group (Figure 1B). RAC values in the control group were quite high and increased over time, while the BPD group had the opposite results (p<0.05) (Figure 1C). Compared with the control group, the BPD group had lower SOD activity and higher MDA levels on the 7th day and the 14th day (p<0.05) (Figure 1D). All of these results were clearly seen on the 14th day, suggesting the successful establishment of the BPD mouse model.
SILENCING XIST ALLEVIATED LUNG DAMAGE IN BPD MICE:
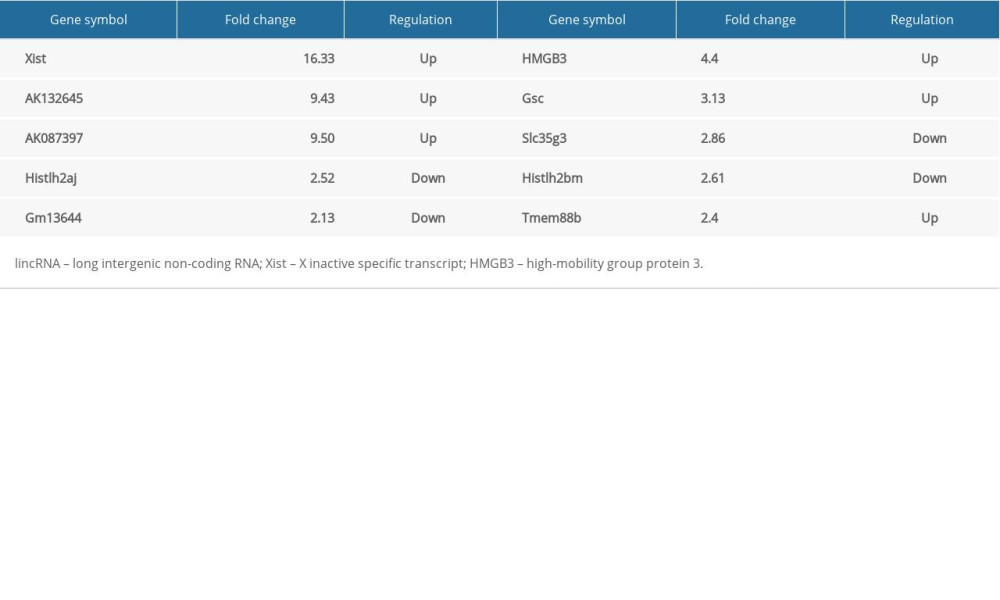
To further explore the mechanism of BPD injury in newborn mice, lncRNA microarray analysis was performed to analyze the differentially expressed lncRNAs. Initially, RNAs from samples of newborn mice from the BPD group and the control group were extracted and then assessed. We found that the A260/A280 value was from 1.8 to 2.0, showing the completeness and purity of the extracted RNA. All the images of microarray analysis hybrid scans were qualified (Supplementary Figure 2). The microarray analysis quality control results confirmed the satisfactory outcome of the sample cRNA production and the expression spectrum microarray analysis quality, and microarray analysis hybrid results indicated a significantly different fold change in lncRNA Xist between the control group and the BPD group (Figure 2A), which was verified by 5 differentially expressed lncRNAs (Table 2) in tissues of the 2 groups. RT-qPCR showed that, compared with the control group, the BPD group had higher Xist expression (p<0.01) (Figure 2B). In the LV-shXist group, poorly expressed Xist (Figure 2C), decreased alveoli interval and alveolar space, increased alveoli number, complete alveoli structure, and improved RAC value were observed (Figure 2D, 2E) with increased SOD activity and decreased MDA level (Figure 2F). According to RT-qPCR and Western blot analysis, in the BPD group, mRNA and protein levels of VEGF were reduced and mRNA and protein levels of collagen I and α-SMA were increased (Figure 2G, 2H). When Xist expression was silenced, these results were the opposite (all p<0.01).
XIST COMPETITIVELY BINDS TO MIR-101-3P TO ACTIVATE HMGB3 IN THE MOUSE MODEL:
We found significantly different fold changes in HMGB3 expression between the control group and the BPD group (Figure 2A), which was verified by 5 mRNAs (Table 2) in tissues of both the control group and the BPD group. RT-qPCR suggested that mRNA expression of HMGB3 in the BPD group was much higher than that in the control group (p<0.01) (Figure 3A).
Starbase was used to predict the specific binding sites in the sequences between Xist and miR-101-3p and between miR-101-3p and HMGB3 (Figure 3B). According to dual-luciferase reporter gene assay, Xist targeted miR-101-3p and miR-101-3p targeted HMGB3 (p<0.01) (Figure 3C). As the results of the RNA pull-down assay showed, Xist was pulled down by biotinylated miR-101-3p-WT while biotinylated miR-101-3p-MUT could not be enriched into Xist (Figure 3D). RT-qPCR found that poorly expressed miR-101-3p in the BPD group was enhanced by silencing Xist (p<0.01) (Figure 3E), resulting in decreased mRNA expression and protein level of HMGB3 (all p<0.01) (Figure 3F, 3G).
OVEREXPRESSION OF MIR-101-3P MITIGATED LUNG DAMAGE IN BPD MICE:
miR-101-3p expression in lung tissues from the agomir NC group was apparently lower than that in the miR-101-3p agomir group. On the basis of silenced Xist, miR-101-3p expression in lung tissues of the BPD mice was lower after silencing miR-101-3p (p<0.05) (Figure 4A). Compared with the agomir NC group, the miR-101-3p agomir group showed reduced alveoli interval and alveolar space, increased alveolar number large RAC value (p<0.05) (Figure 4B, 4C), elevated SOD activity, decreased MDA level (p<0.05) (Figure 4D), increased mRNA and protein levels of VEGF, and decreased mRNA and protein levels of collagen I and α-SMA (all p<0.01) (Figure 4E, 4F). Additionally, silencing miR-101-3p expression in lung tissues with silenced Xist reversed all the results mentioned above.
SILENCED XIST COMPETITIVELY BINDS TO MIR-101-3P TO SUPPRESS HMGB3 AND DOWNREGULATE THE TGF-β1/SMAD3 AXIS IN THE MOUSE MODEL:
ELISA showed that, compared with the Lv-shNC group, the high protein level of TGF-β1 in the BPD group was reduced with silenced Xist (p<0.01) (Figure 5A). Results from RT-qPCR and Western blot analysis showed that the TGF-β1 levels and Smad3 phosphorylation in lung tissues of the BPD group were inhibited after silencing Xist (all p<0.01), while Smad3 protein showed no significant difference (Figure 5B, 5C). Silencing miR-101-3p in the BPD group with silenced Xist promoted mRNA expression and protein level of TGF-β1 and Smad3 phosphorylation (all p<0.01).
Discussion
As the most prevalent respiratory disease in premature infants, BPD mainly arises from an imbalance in lung impairment and repair during lung development [17]. Microarray analysis showed differentially expressed lncRNAs between normal lung tissues and lung tissues with chronic obstructive lung disease [18]. Xist expression was activated in tissues and cell lines of NSCLC patients [13], indicating the deleterious effects of Xist in lung diseases. Among the theories about BPD, little attention has focussed on lncRNA Xist, so we aimed to explore some novel therapies for BPD based on lncRNA Xist. Consequently, our data showed that silenced lncRNA Xist mitigated BPD by regulating miR-101-3p, HGMB3, and the TGF-β1/Smad3 signaling pathway.
Firstly, functional assays showed that in newborn BPD mice, RAC values and SOD activity declined while MDA level increased. As an index in alveolarization assessment, less RAC was identified in BPD lung tissues than that in the normal ones [19]. Chen et al. recently found that SOD activity in BPD mice was decreased with the enhancement of MDA level [20], which is consistent with our findings. In a related research, it was discovered that increased SOD activity could treat lung damage and hypertension in BPD patients [21]. It was reported that enhancement of MDA level triggered hyperoxia-induced oxidative stress, which aggravated lung damage and impeded vascular and pulmonary tissue growth [22]. Microarray analysis showed a significantly different fold change in lncRNA Xist between the healthy newborn mice and the BPD ones. Compared with normal bronchial epithelial cells, NSCLC cells present highly expressed Xist [23]. Then, it was noticed that VEGF expression was increased while expression of collagen I and α-SMA was decreased in mice with silenced Xist. VEGF knockdown may serve as a diagnosis biomarker in premature infants with BPD [24]. Collagen I is mainly localized in bronchi, alveolar septa, and blood vessels, and was overexpressed in patients with BPD, resulting in failure of alveolar septation during BPD progression [25]. According to a recent study, protein levels of α-SMA in lung tissues and blood vessels of BPD mice induced by hyperoxia were far higher than those in normal mice [26]. These experiments strongly suggest that Xist promotes BPD malignancy.
Additionally, results from microarray analysis showed a significantly different fold change in HMGB3 between the control group and the BPD group. Song et al. discovered that HMGB3 was highly expressed in NSCLC, suggesting poor survival rates [27]. Dual-luciferase reporter gene assay unveiled the targeting relations between Xist and miR-101-3p. miR-101-3p expression was reported to be greatly depressed in serum of patients with idiopathic pulmonary fibrosis [28]. In addition, it was reported that miR-101-3p was associated with various radiological features and forced vital capacity in idiopathic pulmonary fibrosis [28]. Additional experiments found that Xist acted as a sponge to miR-101 to worsen cardiac hypertrophy [29]. In our study, we found a targeting relationship between miR-101-3p and HMGB3. In NSCLC, RT-qPCR detected that HMGB3 expression was promoted [30]. Likewise, as the ceRNA of miR-29b, Xist contributed to augmented HMGB1 expression, thereby promoting hepatic stellate cell activation and autophagy and liver fibrogenesis [31]. This means that Xist competitively bound to miR-101-3p and activated HMGB3 to promote BPD progression. The evidence presented above suggests that overexpression of miR-101-3p can mitigate lung damage in BPD mice.
Furthermore, silenced Xist competitively bound to miR-101-3p to suppress HMGB3 and downregulated the TGF-β1/Smad3 axis. TGF-β-related genes and the TGF-β1/Smad3 axis promoted BPD malignancy by participating in inflammation and remodeling of airway and lung development [32]. In prior research, differential TGF-β1 expression was found between normal infants and BPD infants, and TGF-β1 expression was elevated in BPD patients [5]. When the TGF-β1/Smad3 signaling pathway and TGF-β1-related genes were downregulated, TGF-β1-induced airway remodeling was also inhibited, thus BPD progression was significantly palliated [32]. Chang et al. found that silenced Xist inhibits TGF-β-regulated EMT and NSCLC cell growth [33]. From an overall perspective, our data show that silenced Xist helped block BPD progression.
Conclusions
In summary, our study supports that silenced lncRNA Xist alleviates BPD malignancy by competitively binding to miR-101-3p and downregulating the TGF-β1/Smad3 signaling pathway. These results provide a novel approach for BPD treatment. In the future, we will further explore the underlying mechanism of other targets of Xist. More research is needed to find reliable therapeutic targets for BPD. Although the present study was preclinical research, our findings have therapeutic implications for BPD treatment. Our results need to be confirmed in further experiments before they can be effectively applied in clinical practice.
Figures
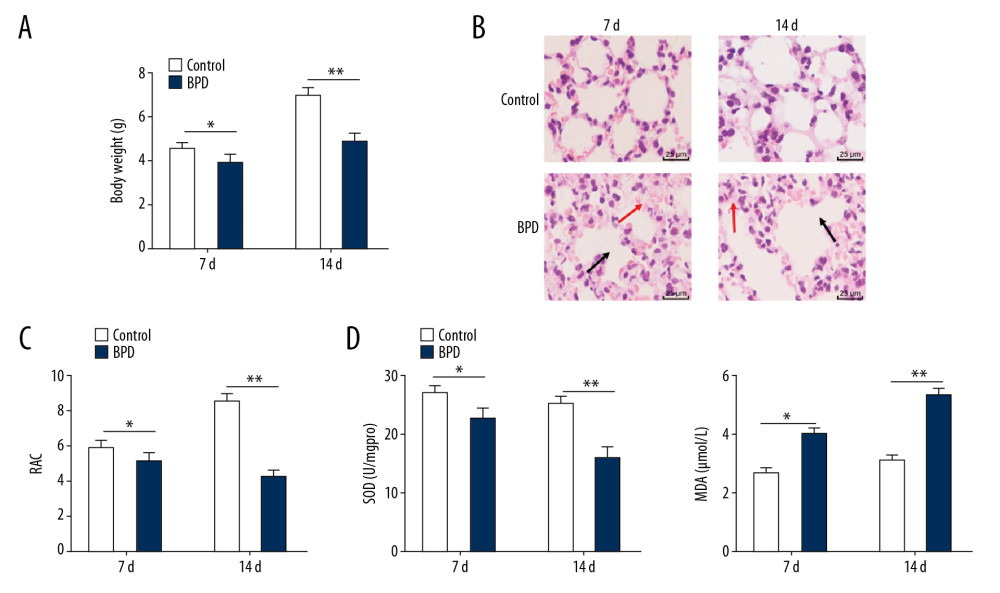 Figure 1. In newborn BPD mice, RAC value and SOD activity were lower and while MDA level was higher. (A) Changes in body weights of BPD newborn mice on the 7th day and the 14th day of BPD modeling, n=5. (B) HE staining showed the variance in lung tissue structure on the 7th day and the 14th day of BPD modeling, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was lower in BPD mice on the 7th day and the 14th day of BPD modeling, n=5. (D) Decreased SOD activity and increased MDA level were found in BPD mice, n=3. Two-way ANOVA and Tukey’s multiple comparisons test were applied to determine statistical significance. * p<0.05, ** p<0.01. BPD – bronchopulmonary dysplasia; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; HE – hematoxylin-eosin; ANOVA – analysis of variance.
Figure 1. In newborn BPD mice, RAC value and SOD activity were lower and while MDA level was higher. (A) Changes in body weights of BPD newborn mice on the 7th day and the 14th day of BPD modeling, n=5. (B) HE staining showed the variance in lung tissue structure on the 7th day and the 14th day of BPD modeling, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was lower in BPD mice on the 7th day and the 14th day of BPD modeling, n=5. (D) Decreased SOD activity and increased MDA level were found in BPD mice, n=3. Two-way ANOVA and Tukey’s multiple comparisons test were applied to determine statistical significance. * p<0.05, ** p<0.01. BPD – bronchopulmonary dysplasia; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; HE – hematoxylin-eosin; ANOVA – analysis of variance. 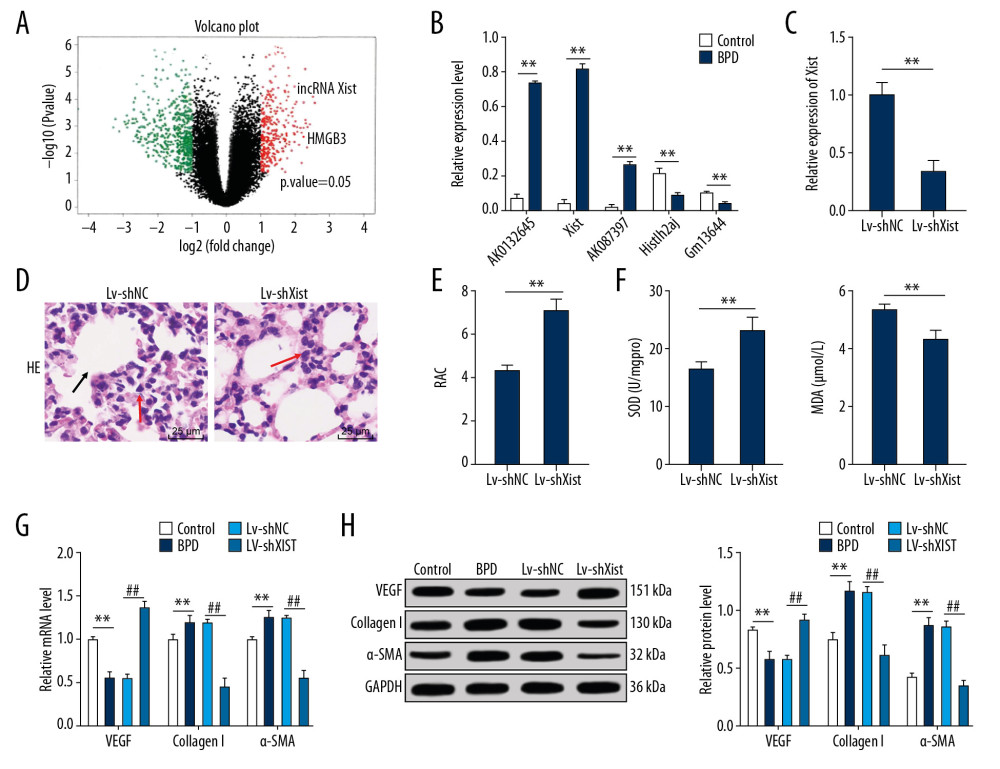 Figure 2. Silenced Xist alleviated lung damage in newborn BPD mice. (A) There was a significantly different fold change in lncRNA expressions between the control group and the BPD group. (B) RT-qPCR showed that the Xist expression was clearly higher in BPD mice than in control mice, n=3. (C) RT-qPCR showed that Xist expression in BPD group was evidently lower than that in the control group with silenced Xist, n=3. (D) HE staining suggested that the lung tissue structure in BPD mice was improved when Xist expression was silenced, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (E) RAC in BPD mice increased with silenced Xist, n=5. (F) ELISA showed that SOD activity was increased while MDA level was decreased in BPD mice with silenced Xist, n=3. (G, H) RT-qPCR and Western blot analysis showed that VEGF mRNA and protein expression levels were reduced while mRNA and protein expression levels of collagen I and α-SMA were increased, n=3. Two-way ANOVA was applied to assess data in panels B, G, and H. Tukey’s multiple comparisons test was used for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. Xist – X inactive specific transcript; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance.
Figure 2. Silenced Xist alleviated lung damage in newborn BPD mice. (A) There was a significantly different fold change in lncRNA expressions between the control group and the BPD group. (B) RT-qPCR showed that the Xist expression was clearly higher in BPD mice than in control mice, n=3. (C) RT-qPCR showed that Xist expression in BPD group was evidently lower than that in the control group with silenced Xist, n=3. (D) HE staining suggested that the lung tissue structure in BPD mice was improved when Xist expression was silenced, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (E) RAC in BPD mice increased with silenced Xist, n=5. (F) ELISA showed that SOD activity was increased while MDA level was decreased in BPD mice with silenced Xist, n=3. (G, H) RT-qPCR and Western blot analysis showed that VEGF mRNA and protein expression levels were reduced while mRNA and protein expression levels of collagen I and α-SMA were increased, n=3. Two-way ANOVA was applied to assess data in panels B, G, and H. Tukey’s multiple comparisons test was used for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. Xist – X inactive specific transcript; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance. 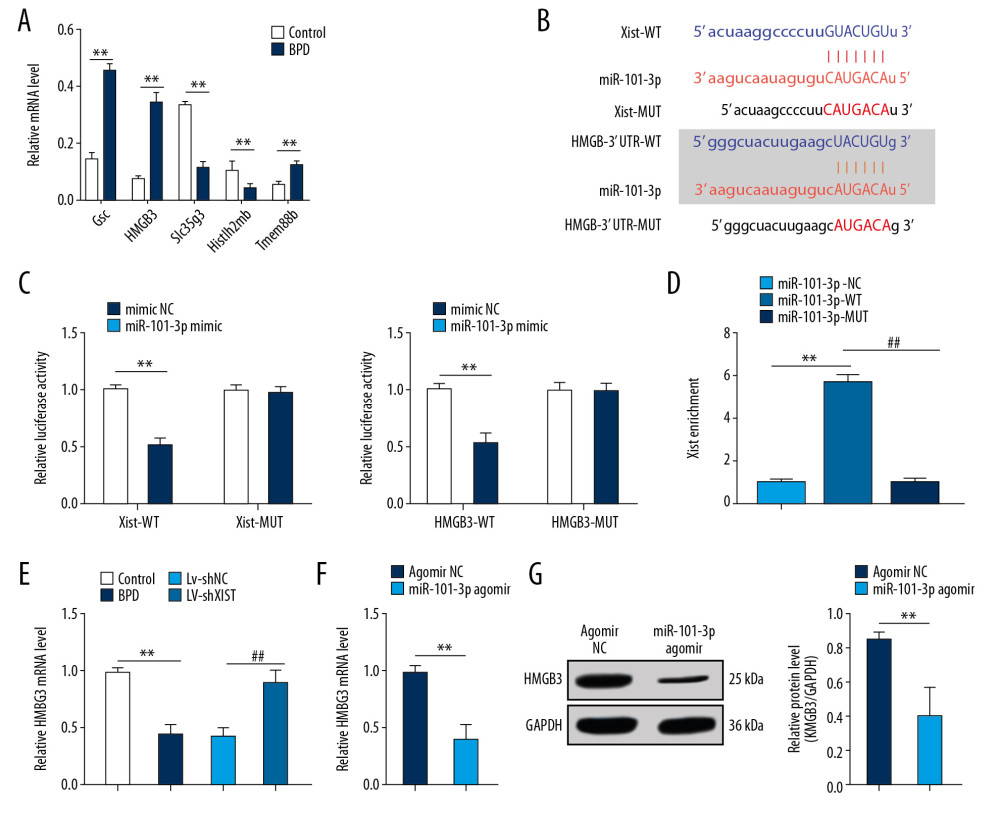 Figure 3. Xist competitively binds to miR-101-3p to regulate HMGB3. (A) RT-qPCR showed that mRNA expression of HMGB3 in the BPD group was evidently higher than that in the control group. (B) Starbase verified the sequence binding sites between Xist and miR-101-3p and between miR-101-3p and HMGB3. (C) Dual-luciferase reporter gene assay verified the targeting relations between Xist and miR-101-3p and between miR-101-3p and HMGB3. (D) RNA pull-down assay further affirmed the targeting relation between Xist and miR-101-3p. (E) RT-qPCR assessed miR-101-3p expression in mice with different treatment. (F, G) RT-qPCR and Western blot analysis showed that mRNA expression and protein level of HMGB3 in BPD mice were decreased with overexpressed miR-101-3p. Two-way ANOVA was used to assess data in panels A and C, one-way ANOVA was used to assess data in panels D and E. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; BPD – bronchopulmonary dysplasia; ANOVA – analysis of variance.
Figure 3. Xist competitively binds to miR-101-3p to regulate HMGB3. (A) RT-qPCR showed that mRNA expression of HMGB3 in the BPD group was evidently higher than that in the control group. (B) Starbase verified the sequence binding sites between Xist and miR-101-3p and between miR-101-3p and HMGB3. (C) Dual-luciferase reporter gene assay verified the targeting relations between Xist and miR-101-3p and between miR-101-3p and HMGB3. (D) RNA pull-down assay further affirmed the targeting relation between Xist and miR-101-3p. (E) RT-qPCR assessed miR-101-3p expression in mice with different treatment. (F, G) RT-qPCR and Western blot analysis showed that mRNA expression and protein level of HMGB3 in BPD mice were decreased with overexpressed miR-101-3p. Two-way ANOVA was used to assess data in panels A and C, one-way ANOVA was used to assess data in panels D and E. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; BPD – bronchopulmonary dysplasia; ANOVA – analysis of variance. 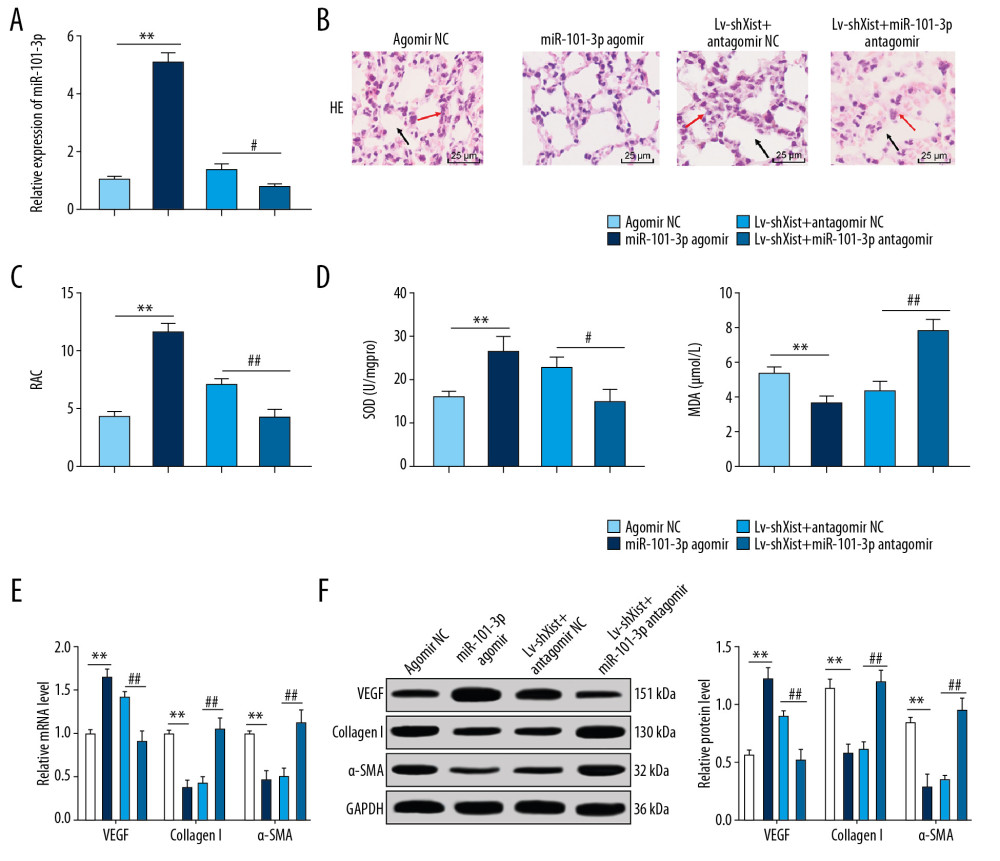 Figure 4. Overexpressed miR-101-3p mitigated lung damage in newborn BPD mice. (A) RT-qPCR showed silencing miR-101-3p in lung tissues reduced miR-101-3p expression in BPD mice, n=3. (B) HE staining indicated the structural changes in lung tissues of BPD mice after silencing miR-101-3p, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was enhanced when miR-101-3p was overexpressed, n=5. (D) ELISA showed improved SOD activity and decreased MDA level with overexpressed miR-101-3p, n=3. (E, F) RT-qPCR and Western blot analysis suggested that mRNA expression and protein level in VEGF were enhanced while those in collagen I and α-SMA were reduced, n=3. Two-way ANOVA was applied to assess data in panels E, F, and one-way ANOVA was applied to assess data in panels A, C, and D. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. miR – microRNA; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance.
Figure 4. Overexpressed miR-101-3p mitigated lung damage in newborn BPD mice. (A) RT-qPCR showed silencing miR-101-3p in lung tissues reduced miR-101-3p expression in BPD mice, n=3. (B) HE staining indicated the structural changes in lung tissues of BPD mice after silencing miR-101-3p, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was enhanced when miR-101-3p was overexpressed, n=5. (D) ELISA showed improved SOD activity and decreased MDA level with overexpressed miR-101-3p, n=3. (E, F) RT-qPCR and Western blot analysis suggested that mRNA expression and protein level in VEGF were enhanced while those in collagen I and α-SMA were reduced, n=3. Two-way ANOVA was applied to assess data in panels E, F, and one-way ANOVA was applied to assess data in panels A, C, and D. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. miR – microRNA; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance. 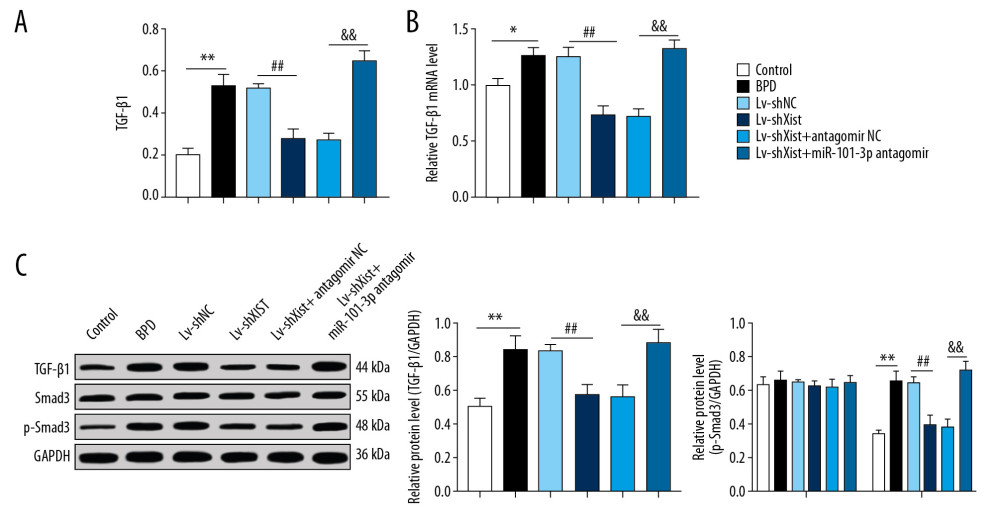 Figure 5. Silenced Xist competitively combines with miR-101-3p to suppress HMGB3 and negatively regulates the TGF-β1/Smad3 axis. (A) ELISA found that the high protein level of TGF-β1 in BPD mice was reduced with silenced Xist. (B) RT-qPCR showed that the highly expressed mRNA of TGF-β1 in lung tissues of newborn BPD mice were inhibited after silencing Xist. (C) Western blot analysis suggested that the high protein level of TGF-β1 and Smad3 phosphorylation in lung tissues of BPD mice were inhibited after silencing Xist. One-way ANOVA was performed to determine data in panels A–C, and two-way ANOVA was performed to determine data in panel D, Tukey’s multiple comparisons test was applied for post hoc test. * p<0.01, ** p<0.01, ## p<0.01, && p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; TGF-β1 – transforming growth factor-beta 1; Smad3 – drosophila mothers against decapentaplegic 3; ELISA – enzyme-linked immunosorbent assay; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; ANOVA – analysis of variance.
Figure 5. Silenced Xist competitively combines with miR-101-3p to suppress HMGB3 and negatively regulates the TGF-β1/Smad3 axis. (A) ELISA found that the high protein level of TGF-β1 in BPD mice was reduced with silenced Xist. (B) RT-qPCR showed that the highly expressed mRNA of TGF-β1 in lung tissues of newborn BPD mice were inhibited after silencing Xist. (C) Western blot analysis suggested that the high protein level of TGF-β1 and Smad3 phosphorylation in lung tissues of BPD mice were inhibited after silencing Xist. One-way ANOVA was performed to determine data in panels A–C, and two-way ANOVA was performed to determine data in panel D, Tukey’s multiple comparisons test was applied for post hoc test. * p<0.01, ** p<0.01, ## p<0.01, && p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; TGF-β1 – transforming growth factor-beta 1; Smad3 – drosophila mothers against decapentaplegic 3; ELISA – enzyme-linked immunosorbent assay; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; ANOVA – analysis of variance. 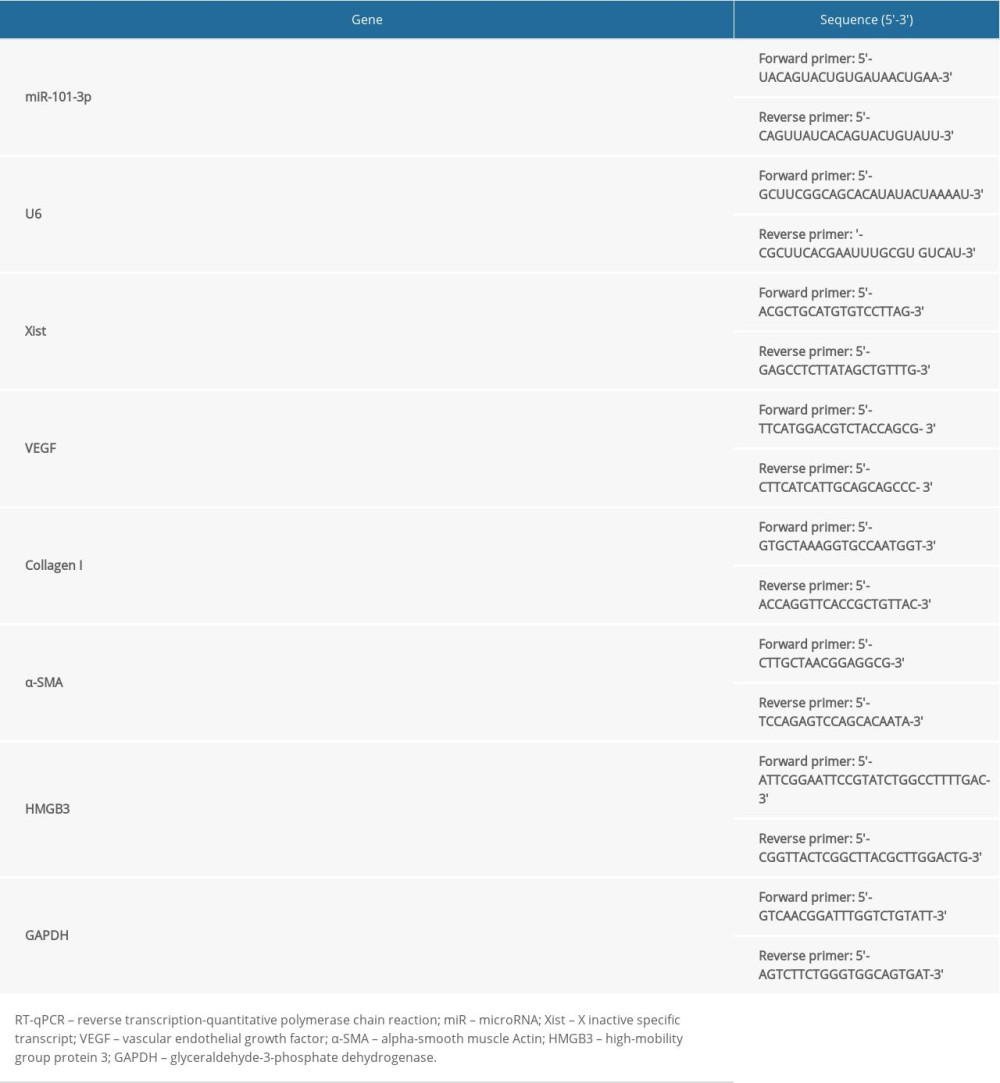
References
1. Stoecklin B, Simpson SJ, Pillow JJ, Bronchopulmonary dysplasia: Rationale for a pathophysiological rather than treatment-based approach to diagnosis: Paediatr Respir Rev, 2019; 32; 91-97
2. Pasha AB, Chen XQ, Zhou GP, Bronchopulmonary dysplasia: Pathogenesis and treatment: Exp Ther Med, 2018; 16(6); 4315-21
3. Berger J, Bhandari V, Animal models of bronchopulmonary dysplasia. The term mouse models: Am J Physiol Lung Cell Mol Physiol, 2014; 307(12); L936-47
4. Wang J, Dong W, Oxidative stress and bronchopulmonary dysplasia: Gene, 2018; 678; 177-83
5. Aly H, Mansi Y, Ez El Din Z, Mesenchymal stromal cells and TGF-beta1 in tracheal aspirate of premature infants: Early predictors for bronchopulmonary dysplasia?: J Perinat Med, 2019; 47(4); 470-77
6. Hwang JS, Rehan VK, Recent advances in bronchopulmonary dysplasia: Pathophysiology, prevention, and treatment: Lung, 2018; 196(2); 129-38
7. Nardiello C, Mizikova I, Morty RE, Looking ahead: Where to next for animal models of bronchopulmonary dysplasia?: Cell Tissue Res, 2017; 36(3); 457-68
8. Yin J, Wang X, Zhang L, Peptidome analysis of lung tissues from a hyperoxia-induced bronchopulmonary dysplasia mouse model: Insights into the pathophysiological process of bronchopulmonary dysplasia: J Cell Physiol, 2018; 233(10); 7101-12
9. Guo ZH, You ZH, Wang YB, A learning-based method for LncRNA-disease association identification combing similarity information and rotation forest: iScience, 2019; 19; 786-95
10. Bao TP, Wu R, Cheng HP, Differential expression of long non-coding RNAs in hyperoxia-induced bronchopulmonary dysplasia: Cell Biochem Funct, 2016; 34(5); 299-309
11. Wang J, Yin J, Wang X, Changing expression profiles of mRNA, lncRNA, circRNA, and miRNA in lung tissue reveal the pathophysiological of bronchopulmonary dysplasia (BPD) in mouse model: J Cell Biochem, 2019; 120(6); 9369-80
12. Tang Y, He R, An J, lncRNA XIST interacts with miR-140 to modulate lung cancer growth by targeting iASPP: Oncol Rep, 2017; 38(2); 941-48
13. Wang H, Shen Q, Zhang X, The long non-coding RNA XIST controls non-small cell lung cancer proliferation and invasion by modulating miR-186-5p: Cell Physiol Biochem, 2017; 41(6); 2221-29
14. Jiang H, Zhang H, Hu X, Li W, Knockdown of long non-coding RNA XIST inhibits cell viability and invasion by regulating miR-137/PXN axis in non-small cell lung cancer: Int J Biol Macromol, 2018; 111; 623-31
15. Vishnoi A, Rani S, MiRNA biogenesis and regulation of diseases: An overview: Methods Mol Biol, 2017; 1509; 1-10
16. Yang Y, Qiu J, Kan Q, MicroRNA expression profiling studies on bronchopulmonary dysplasia: A systematic review and meta-analysis: Genet Mol Res, 2013; 12(4); 5195-206
17. Kalikkot Thekkeveedu R, Guaman MC, Shivanna B, Bronchopulmonary dysplasia: A review of pathogenesis and pathophysiology: Respir Med, 2017; 132; 170-77
18. Zhang J, Zhu Y, Wang R, Long noncoding RNAs in respiratory diseases: Histol Histopathol, 2018; 33(8); 747-56
19. Tokuriki S, Igarashi A, Okuno T, Treatment with geranylgeranylacetone induces heat shock protein 70 and attenuates neonatal hyperoxic lung injury in a model of bronchopulmonary dysplasia: Lung, 2017; 195(4); 469-76
20. Chen X, Zhang X, Pan J, Effect of montelukast on bronchopulmonary dysplasia (BPD) and related mechanisms: Med Sci Monit, 2019; 25; 1886-93
21. Sherlock LG, Trumpie A, Hernandez-Lagunas L, Redistribution of extracellular superoxide dismutase causes neonatal pulmonary vascular remodeling and PH but protects against experimental bronchopulmonary dysplasia: Antioxidants (Basel), 2018; 7(3) pii: E42
22. Maturu P, Wei-Liang Y, Androutsopoulos VP, Quercetin attenuates the hyperoxic lung injury in neonatal mice: Implications for bronchopulmonary dysplasia (BPD): Food Chem Toxicol, 2018; 114; 23-33
23. Zhang YL, Li XB, Hou YX, The lncRNA XIST exhibits oncogenic properties via regulation of miR-449a and Bcl-2 in human non-small cell lung cancer: Acta Pharmacol Sin, 2017; 38(3); 371-81
24. Taglauer E, Abman SH, Keller RL, Recent advances in antenatal factors predisposing to bronchopulmonary dysplasia: Semin Perinatol, 2018; 42(7); 413-24
25. Mizikova I, Morty RE, The extracellular matrix in bronchopulmonary dysplasia: Target and source: Front Med (Lausanne), 2015; 2; 91
26. Trittmann JK, Velten M, Heyob KM, Arginase and alpha-smooth muscle actin induction after hyperoxic exposure in a mouse model of bronchopulmonary dysplasia: Clin Exp Pharmacol Physiol, 2018; 45(6); 556-62
27. Song N, Wang B, Feng G, Knockdown of high mobility group box 3 impairs cell viability and colony formation but increases apoptosis in A549 human non-small cell lung cancer cells: Oncol Lett, 2019; 17(3); 2937-45
28. Li P, Li J, Chen T, Expression analysis of serum microRNAs in idiopathic pulmonary fibrosis: Int J Mol Med, 2014; 33(6); 1554-62
29. Xiao L, Gu Y, Sun Y, The long noncoding RNA XIST regulates cardiac hypertrophy by targeting miR-101: J Cell Physiol, 2019; 234(8); 13680-92
30. Shi J, Wang H, Feng W, Long non-coding RNA HOTTIP promotes hypoxia-induced glycolysis through targeting miR-615-3p/HMGB3 axis in non-small cell lung cancer cells: Eur J Pharmacol, 2019; 862; 172615
31. Xie ZY, Wang FF, Xiao ZH, Long noncoding RNA XIST enhances ethanol-induced hepatic stellate cells autophagy and activation via miR-29b/HMGB1 axis: IUBMB Life, 2019; 71(12); 1962-72
32. Kunzmann S, Ottensmeier B, Speer CP, Fehrholz M, Effect of progesterone on Smad signaling and TGF-beta/Smad-regulated genes in lung epithelial cells: PLoS One, 2018; 13(7); e0200661
33. Li C, Wan L, Liu Z, Long non-coding RNA XIST promotes TGF-beta-induced epithelial-mesenchymal transition by regulating miR-367/141-ZEB2 axis in non-small-cell lung cancer: Cancer Lett, 2018; 418; 185-95
Figures
Figure 1. In newborn BPD mice, RAC value and SOD activity were lower and while MDA level was higher. (A) Changes in body weights of BPD newborn mice on the 7th day and the 14th day of BPD modeling, n=5. (B) HE staining showed the variance in lung tissue structure on the 7th day and the 14th day of BPD modeling, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was lower in BPD mice on the 7th day and the 14th day of BPD modeling, n=5. (D) Decreased SOD activity and increased MDA level were found in BPD mice, n=3. Two-way ANOVA and Tukey’s multiple comparisons test were applied to determine statistical significance. * p<0.05, ** p<0.01. BPD – bronchopulmonary dysplasia; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; HE – hematoxylin-eosin; ANOVA – analysis of variance.Figure 2. Silenced Xist alleviated lung damage in newborn BPD mice. (A) There was a significantly different fold change in lncRNA expressions between the control group and the BPD group. (B) RT-qPCR showed that the Xist expression was clearly higher in BPD mice than in control mice, n=3. (C) RT-qPCR showed that Xist expression in BPD group was evidently lower than that in the control group with silenced Xist, n=3. (D) HE staining suggested that the lung tissue structure in BPD mice was improved when Xist expression was silenced, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (E) RAC in BPD mice increased with silenced Xist, n=5. (F) ELISA showed that SOD activity was increased while MDA level was decreased in BPD mice with silenced Xist, n=3. (G, H) RT-qPCR and Western blot analysis showed that VEGF mRNA and protein expression levels were reduced while mRNA and protein expression levels of collagen I and α-SMA were increased, n=3. Two-way ANOVA was applied to assess data in panels B, G, and H. Tukey’s multiple comparisons test was used for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. Xist – X inactive specific transcript; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance.Figure 3. Xist competitively binds to miR-101-3p to regulate HMGB3. (A) RT-qPCR showed that mRNA expression of HMGB3 in the BPD group was evidently higher than that in the control group. (B) Starbase verified the sequence binding sites between Xist and miR-101-3p and between miR-101-3p and HMGB3. (C) Dual-luciferase reporter gene assay verified the targeting relations between Xist and miR-101-3p and between miR-101-3p and HMGB3. (D) RNA pull-down assay further affirmed the targeting relation between Xist and miR-101-3p. (E) RT-qPCR assessed miR-101-3p expression in mice with different treatment. (F, G) RT-qPCR and Western blot analysis showed that mRNA expression and protein level of HMGB3 in BPD mice were decreased with overexpressed miR-101-3p. Two-way ANOVA was used to assess data in panels A and C, one-way ANOVA was used to assess data in panels D and E. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; BPD – bronchopulmonary dysplasia; ANOVA – analysis of variance.Figure 4. Overexpressed miR-101-3p mitigated lung damage in newborn BPD mice. (A) RT-qPCR showed silencing miR-101-3p in lung tissues reduced miR-101-3p expression in BPD mice, n=3. (B) HE staining indicated the structural changes in lung tissues of BPD mice after silencing miR-101-3p, with the black arrows indicating alveolar fusion and the red ones indicating alveolar septum, ×400, n=5. (C) RAC was enhanced when miR-101-3p was overexpressed, n=5. (D) ELISA showed improved SOD activity and decreased MDA level with overexpressed miR-101-3p, n=3. (E, F) RT-qPCR and Western blot analysis suggested that mRNA expression and protein level in VEGF were enhanced while those in collagen I and α-SMA were reduced, n=3. Two-way ANOVA was applied to assess data in panels E, F, and one-way ANOVA was applied to assess data in panels A, C, and D. Tukey’s multiple comparisons test was applied for post hoc test. The t test was used for analyzing data in remaining panels. ** p<0.01, ## p<0.01. miR – microRNA; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; HE – hematoxylin-eosin; RAC – radial alveolar counts; SOD – superoxide dismutase; MDA – malondialdehyde; VEGF – vascular endothelial growth factor; α-SMA – alpha-smooth muscle Actin; ANOVA – analysis of variance.Figure 5. Silenced Xist competitively combines with miR-101-3p to suppress HMGB3 and negatively regulates the TGF-β1/Smad3 axis. (A) ELISA found that the high protein level of TGF-β1 in BPD mice was reduced with silenced Xist. (B) RT-qPCR showed that the highly expressed mRNA of TGF-β1 in lung tissues of newborn BPD mice were inhibited after silencing Xist. (C) Western blot analysis suggested that the high protein level of TGF-β1 and Smad3 phosphorylation in lung tissues of BPD mice were inhibited after silencing Xist. One-way ANOVA was performed to determine data in panels A–C, and two-way ANOVA was performed to determine data in panel D, Tukey’s multiple comparisons test was applied for post hoc test. * p<0.01, ** p<0.01, ## p<0.01, && p<0.01, n=3. Xist – X inactive specific transcript; miR – microRNA; HMGB3 – high-mobility group protein B3; TGF-β1 – transforming growth factor-beta 1; Smad3 – drosophila mothers against decapentaplegic 3; ELISA – enzyme-linked immunosorbent assay; BPD – bronchopulmonary dysplasia; RT-qPCR – reverse transcription-quantitative polymerase chain reaction; ANOVA – analysis of variance. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387