25 August 2020: Lab/In Vitro Research
Dioscin Attenuates Interleukin 1β (IL-1β)-Induced Catabolism and Apoptosis via Modulating the Toll-Like Receptor 4 (TLR4)/Nuclear Factor kappa B (NF-κB) Signaling in Human Nucleus Pulposus Cells
Longhui Wang1DEF, Yuntao Gu1BE, Hai Zhao1CEF, Rong Chen1BD, Wensheng Chen1BF, Hao Qi1BE, Weisong Gao1ABG*DOI: 10.12659/MSM.923386
Med Sci Monit 2020; 26:e923386






Abstract
BACKGROUND: Nucleus pulposus (NP) cell dysfunction and apoptosis contribute to disc degeneration. Dioscin, a natural steroid saponin, has been demonstrated to have anti-inflammatory, antiapoptotic, and antioxidative effects in various diseases. However, little is known about the roles of dioscin in intervertebral disc degeneration.
MATERIAL AND METHODS: To evaluate the roles of dioscin in disc degeneration and its specific mechanism, human NP cells were incubated with IL-1β and various concentrations of dioscin. Cell viability, extracellular matrix protein expression, catabolic factors, degree of apoptosis, inflammatory factors, and related signaling pathways were evaluated by western blotting, fluorescence immunostaining, TUNEL staining, and reverse transcription PCR.
RESULTS: Dioscin inhibited IL-1β-activated apoptotic signaling and catabolic activity in NP cells. Dioscin suppressed TLR4/NF-0κB signaling, and attenuated the level of inflammatory mediators (IL-6, TNF-α) in IL-1β-stimulated human NP cells.
CONCLUSIONS: Our work provides the first evidence that dioscin attenuates IL-1β-activated inflammation and catabolic activity in human NP cells through inhibiting the TLR4/NF-κB pathway, indicating that dioscin is a new potential candidate for clinical therapy to attenuate disc degeneration.
Keywords: intervertebral disc degeneration, Diosgenin, Inflammation Mediators, nucleus pulposus
Background
The intervertebral disc is an important component of the spinal structure, which absorbs, alleviates, and transmits loads on the human body and increases the mobility of the spine [1]. However, disc decompensation under certain conditions can lead to progressive destruction of its structure, resulting in disc herniation, discogenic pain, and other disc degeneration-related diseases (e.g., intervertebral disc disease [IDD]), which impose a serious burden on patients’ lives as well as socioeconomic development. It has been estimated that more than half of the population will suffer from low back pain at different stages of life [2], and more than 40% of cases of low back pain are caused by disc degeneration [3]. Research on disc degeneration has mainly focused on the changes in metabolites during the stress process of the intervertebral disc itself. It is believed that the structure of the intervertebral disc can be destroyed under stimulation by stress factors, such as abnormal stress and nutritional dysfunction, which can lead to cell metabolic disorders, inflammation, apoptosis, autophagy, and other phenomena, eventually leading to disc degeneration [4,5]. However, the pathophysiological mechanism of IDD is unclear, necessitating further research to develop effective preventive and treatment strategies.
Various pathological events, such as mechanical stress, structural deformities, genetic factors, infection, and smoking, activate pro-inflammatory signaling pathways in disc cells. These initializing triggers cause morphological changes in the disc tissues, thereby triggering further inflammation and degenerative processes. Nucleus pulposus (NP) cells are present in a gelatinous extracellular matrix (ECM) containing collagen II and proteoglycan, which are essential for resistance to compressive axial force of the spine [6,7]. In healthy NP tissue, the NP cell maintains the metabolic balance of ECM, including aggrecan and collagen, with long half-lives [8]. Studies have demonstrated that matrix metalloproteinases (MMPs) and a disintegrin and metalloproteinase with thrombospondin type I motifs (ADAMTSs) disrupt the balance of ECM metabolism during the pathological process of disc degeneration.
Various pro-inflammatory mediators containing tumor necrosis factor (TNF)-α and interleukin (IL)-1β have been shown to be significantly up-regulated in degenerative disc tissue in humans [9,10]. TNF-α and IL-6, essential pro-inflammatory factors, have been shown to be closely related to the progression of IDD [11]. TNF-α can trigger inflammation, leading to neural swelling and neuropathic pain, and aggravate cell apoptosis due to its cytotoxic effects in humans with lower lumbar pain. IL-1β, the most studied proinflammatory factor, has been shown to trigger the level of various proinflammatory mediators, including TNF-α, IL-6, and several matrix-degrading enzymes, disrupting the balance of ECM metabolism and impairing its turnover in the intervertebral discs [12]. Several molecular pathways, such as nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) signaling, have been shown to be major moderators of IL-1β-induced inflammation and catabolism [13].
Dioscin, a natural steroid saponin, is extracted from several herbal medicines [14]. Pharmacological studies have suggested that dioscin has anti-inflammatory, anti-apoptotic, and anti-oxidative effects in various diseases [15–19]. For instance, dioscin was shown to attenuate dimethylnitrosamine-induced acute liver injury by inhibiting apoptosis via reductions in the expression of proapoptotic proteins [16]. Dioscin inhibited hepatocellular carcinoma via regulation of apoptosis, autophagy, and DNA damage [20] and it attenuated intestinal ischemia/reperfusion injury through regulating miR-351-5p-related oxidative stress, inflammation, and apoptosis [21,22]. Dioscin markedly prevented non-alcoholic fatty liver disease via regulating lipid metabolism by activating SIRT1/AMPK signaling [23]. Dioscin inhibited systemic inflammatory response syndromevia via adjusting the TLR2/NF κb signaling pathway [24]. Dioscin was also reported to attenuate cerebral ischemia/reperfusion injury via inhibiting TLR4 signaling [25].
However, little is known about the potential effects of dioscin on IDD. Therefore, this study aimed to explore the potential effects of dioscin in IL-1β-treated NP cells and its specific mechanism of action.
Material and methods
CHEMICALS AND MATERIALS:
Dioscin (Figure 1) with a purity >99% was purchased from Shanghai Tauto Biochemical Technology Co., Ltd. (Shanghai, China). IL-1β was acquired from R&D Systems (St. Paul, MN, USA). Antibodies were acquired from Santa Cruz (TLR4, P65, and IκBα; Santa Cruz, CA, USA) and Cell Signaling Technology (collagen II, cleaved caspase-3, caspase-9, Bcl-2, and β-actin; Beverly, MA, USA). Other reagents not mentioned here were acquired from Sigma-Aldrich.
PRIMARY HUMAN NP CELL CULTURE AND TREATMENT:
Human NP tissue collection conformed to the guidelines of the Ethics Committee of the Second Affiliated Hospital of Hainan Medical University. Informed consent for disc tissue collection was acquired from patients or their relatives. The protocol of human NP cell culture was performed as described previously [26]. NP cells at passage 1–2 were prepared for reverse transcription PCR, immunofluorescence assay, and western blotting. The cells were planted into 6-well plates. After 2 days, the samples were treated with dioscin (1, 10, 100, 500 ng/mL) supplemented with IL-1β (10 ng/mL) for 1 day, and NP cells were then used for the experiments.
VIABILITY ASSAY:
The NP cells were seeded and cultured in 96-well plates (5–6×103 cells/well) for 1 day and incubated with the various concentrations of dioscin with or without IL-1β (10 ng/mL). After incubation for 1 day, CCK-8 solution (10μL) was added to each well for 2 h. The absorbance of samples was tested at 450 nm by a spectrophotometer.
IMMUNOFLUORESCENCE ASSAY:
NP cells were seeded and cultured on coverslips, treated with 4% paraformaldehyde for 15 min, and then treated in 0.5% Triton X-100 for 10 min. Following incubation in blocking buffer containing 5% BSA for 30 min, samples were treated with the primary antibody at 4°C. The following day, samples were treated with the secondary antibody for 1 h at 37°C. Sample were sealed with VECTASHIELD (Vector Laboratories, Burlingame, CA, USA) including DAPI. The immunostained samples were imaged by laser scanning confocal microscopy.
WESTERN BLOTTING:
All protein samples were isolated using cell lysis buffer. The concentration of total protein was tested with a BCA protein assay kit. The protein specimens were added onto SDS gel electrophoresis and transferred onto PVDF membrane. After blocking nonspecific binding sites, the membranes were immunoblotted with primary antibodies. The following day, samples were treated with the secondary antibody for 60 min at 37°C. Band intensity was detected using the ChemiDoc XRS+ Imaging System.
TUNEL ASSAY:
After treatment, NP cells were incubated with 4% paraformaldehyde, and then treated in 0.5% Triton X-100 for 10 min. Apoptotic NP cells were stained with an In Situ Cell Death Detection Kit (Roche, Basel, Switzerland) according to the manufacturer’s instructions, and DAPI. The degree of apoptotic level in each group was determined by laser scanning confocal microscopy.
REVERSE TRANSCRIPTION PCR ASSAY:
Total RNA of NP cells was extracted using the TRIzol method. cDNAs were synthetized by reverse transcription using oligo (dT) with RNA samples. cDNAs were amplified with a SYBR Green PCR kit. The primers of the targeted genes are listed in Table 1. mRNA expression was quantified using the ddCt method.
STATISTICAL ANALYSIS:
Results were analyzed as means±standard deviations (SD). Differences among groups were tested by one-way analysis of variance and Tukey’s post hoc test.
Results
EFFECTS OF DIOSCIN ON IL-1β-ACTIVATED CELL VIABILITY IN HUMAN NP CELLS:
We first examined the safe dose range of dioscin that did not adversely affect the NP cells viability using the CCK-8 method. Dioscin exhibited no effects on cell viability at doses of 1 to 1000 ng/mL (Figure 2A). NP cells were incubated with IL-1β and different concentrations of dioscin (1, 10, 100, and 500 ng/mL). Dioscin showed a dose-dependent inhibitory effect on IL-1β-induced cell viability (Figure 2B).
EFFECTS OF DIOSCIN ON IL-1β-ACTIVATED APOPTOTIC SIGNALING IN HUMAN NP CELLS:
To clarify the molecular events related to the roles of dioscin in IL-1β-activated apoptotic activity, the level of cleaved caspase-3, caspase-9, and Bcl-2 were determined. Treatment with IL-1β markedly up-regulated the level of cleaved caspase-3 and caspase-9, and down-regulated Bcl-2 expression. However, dioscin dose-dependently attenuated these IL-1β-induced alterations in level of cleaved caspase-3, caspase-9, and Bcl-2 (Figure 3A–3D). Furthermore, the results of TUNEL staining showed similar anti-apoptotic roles of dioscin in IL-1β-treated NP cells as was observed in untreated controls (Figure 3E, 3F). The immunofluorescence data of cleaved caspase-3 were consistent with the above results (Figure 3G). These results suggested that dioscin alleviated IL-1β-activated apoptotic activity in human NP cells.
DIOSCIN SUPPRESSED IL-1β-ACTIVATED CATABOLIC ACTIVITY IN HUMAN NP CELLS:
Catabolic enzymes including MMP3 and MMP13 together with ADAMTS4 and ADAMTS5 are closely related to the ECM catabolism of NP cells. Thus, we measured these catabolic factors by PCR to assess the catabolic activity of the NP cells. As shown in Figure 4, the expressions of ADAMTS4 and ADAMTS5 were sharply up-regulated in the IL-1β group and markedly reversed by dioscin. Similarly, the mRNA expressions of MMP3 and MMP13, which were increased in the IL-1β group, were markedly attenuated by dioscin. These results showed that dioscin treatment inhibited IL-1β-activated catabolic activity in human NP cells.
DIOSCIN ATTENUATED ECM DEGRADATION IN IL-1β-STIMULATED HUMAN NP CELLS:
The levels of proteins and mRNAs involved in ECM degradation were measured by PCR and immunofluorescence assay. The levels of collagen II and aggrecan, major components of the ECM, were markedly decreased by IL-1β (Figure 5A, 5B). However, dioscin-treated NP cells showed higher levels of collagen II and aggrecan mRNA compared to the IL-1-βtreated cells. The results of immunofluorescence staining for collagen II were consistent with the results of the PCR assays (Figure 5C). These data suggested that dioscin treatment preserved ECM protein in IL-1β-induced NP cells.
DIOSCIN INHIBITED IL-1β-INDUCED RELEASE OF PRO-INFLAMMATORY MEDIATORS BY REGULATION OF NF-κB SIGNALING IN HUMAN NP CELLS:
There is accumulating evidence that inflammation is closely related to disc degeneration. Therefore, we evaluated the mRNA level of the inflammatory factors IL-6 and TNF-α. The PCR results were similar to previous results in that dioscin treatment markedly down-regulated the expressions of IL-6 and TNF-α in IL-1β-treated cells (Figure 6A, 6B). NF-κB activity is critical for inflammation. Therefore, we measured the effects of dioscin on NF-κB activation in the human NP cells. NF-κB protein expression in the dioscin treated groups was markedly reduced compared to that in the IL-1β-treated group (Figure 6C, 6E). In addition, IL-1β-induced degradation of inhibitor of kappa B (IκBα) was evaluated in the human NP cells; treatment with dioscin significantly inhibited IκBα degradation in IL-1β-stimulated human NP cells (Figure 6C, 6F). To explore the effects of the TLR4 pathway on the regulation of NF-κB signaling, the expression of TLR4 was evaluated. The results suggested that TLR4 level was markedly up-regulated in the IL-1β treatment group compared to control NP cells, and dioscin treatment reversed these effects (Figure 6C, 6D). Taken together, these observations indicated that dioscin treatment inhibited inflammation via regulation of TLR4/NF-κB signaling.
DIOSCIN INHIBITED NF-κB TRANSLOCATION TO THE NUCLEUS IN HUMAN NP CELLS:
Immunofluorescence assay showed that overexpressed p65 was translocated into the nucleus in NP cells exposed to IL-1β (Figure 7). However, treatment with dioscin significantly suppressed nuclear translocation of p65 in the IL-1β-treated group (Figure 7).
Discussion
Low back pain causes physical disability, and more than 80% of adults worldwide have suffered from low back pain [27]. IDD has been reported to be one of the major reasons for low back pain [28]. Senescence, metabolic imbalance, mechanical stress, and inflammation are involved in the pathogenesis of IDD [29,30]. However, the specific pathological mechanism underlying IDD has not been fully elucidated. The disc tissue consists of the central NP tissue, peripheral annulus fibrosus tissue, and cartilage endplate. It has been demonstrated that NP cells exhibit a degenerative phenotype in the early stage of IDD [31].
Studies have suggested that inflammation and apoptosis are 2 essential characteristics in NP tissue during the pathological process of IDD [32,33]. In healthy NP tissue, the NP cells maintain the metabolic balance of ECM, including aggrecan and collagen, with long half-lives [8]. A recent study showed that apoptosis of NP cells is closely related to ECM degradation [34]. Furthermore, inflammatory cytokines (IL-1β and TNF-α) attract death-related signaling complexes through interaction with their specific ligands, and then initiate apoptotic signaling, contributing to DNA fragmentation [35]. It was reported that dioscin protected H9c2 cells against hypoxia/reoxygenation injury by regulating mitochondrial apoptotic signaling via inhibition of oxidative stress [36]. Dioscin was also shown to attenuate dimethylnitrosamine-stimulated acute liver injury via suppression of apoptotic activity by decreasing the level of FasL, Fas, p53, Bak, caspase-3, and caspase-9 [16]. Our results suggested that IL-1β promotes NP cell apoptosis, which is partly inhibited by dioscin, suggesting that dioscin exerts an anti-apoptotic effect on NP cells in an inflammatory environment.
It has been reported that the excessive inflammatory response in disc NP tissue is closely related to an increase in the degree of apoptosis of NP cells [37,38]. Therefore, inhibition of the inflammatory response may contribute to the prevention of NP cell apoptosis and reverse degenerative disc changes. TLR signaling pathways were demonstrated to enhance the expression of inflammatory mediators, such as TNF-α, IL-1, and IL-6, contributing to inflammatory responses [39]. TLR4 was shown to be over-expressed in cartilage during the process of osteoarthritis and to play essential roles in cartilage degeneration [40]. Similarly,
Conclusions
TLR4/NF-κB signaling was suggested to be related to the mechanisms of action of dioscin against IL-β-activated inflammatory response, apoptosis, and catabolic activity in human NP cells. Our results provide the first evidence for the pharmacological effects and mechanisms of action of dioscin, which should be considered as a potential new candidate for clinical therapy to attenuate disc degeneration.
Figures
 Figure 1. The molecular structure of dioscin.
Figure 1. The molecular structure of dioscin. 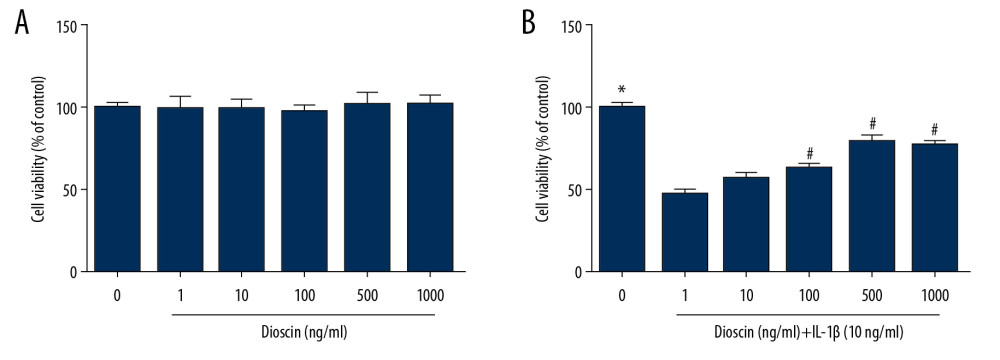 Figure 2. Effects of dioscin on IL-1β-activated cell viability in human NP cells. (A) The cytotoxic effects of dioscin on human NP cells were examined at different concentrations by the CCK-8 method. (B) CCK-8 analysis of dioscin-treated human NP cells stimulated by IL-1β. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.
Figure 2. Effects of dioscin on IL-1β-activated cell viability in human NP cells. (A) The cytotoxic effects of dioscin on human NP cells were examined at different concentrations by the CCK-8 method. (B) CCK-8 analysis of dioscin-treated human NP cells stimulated by IL-1β. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5. 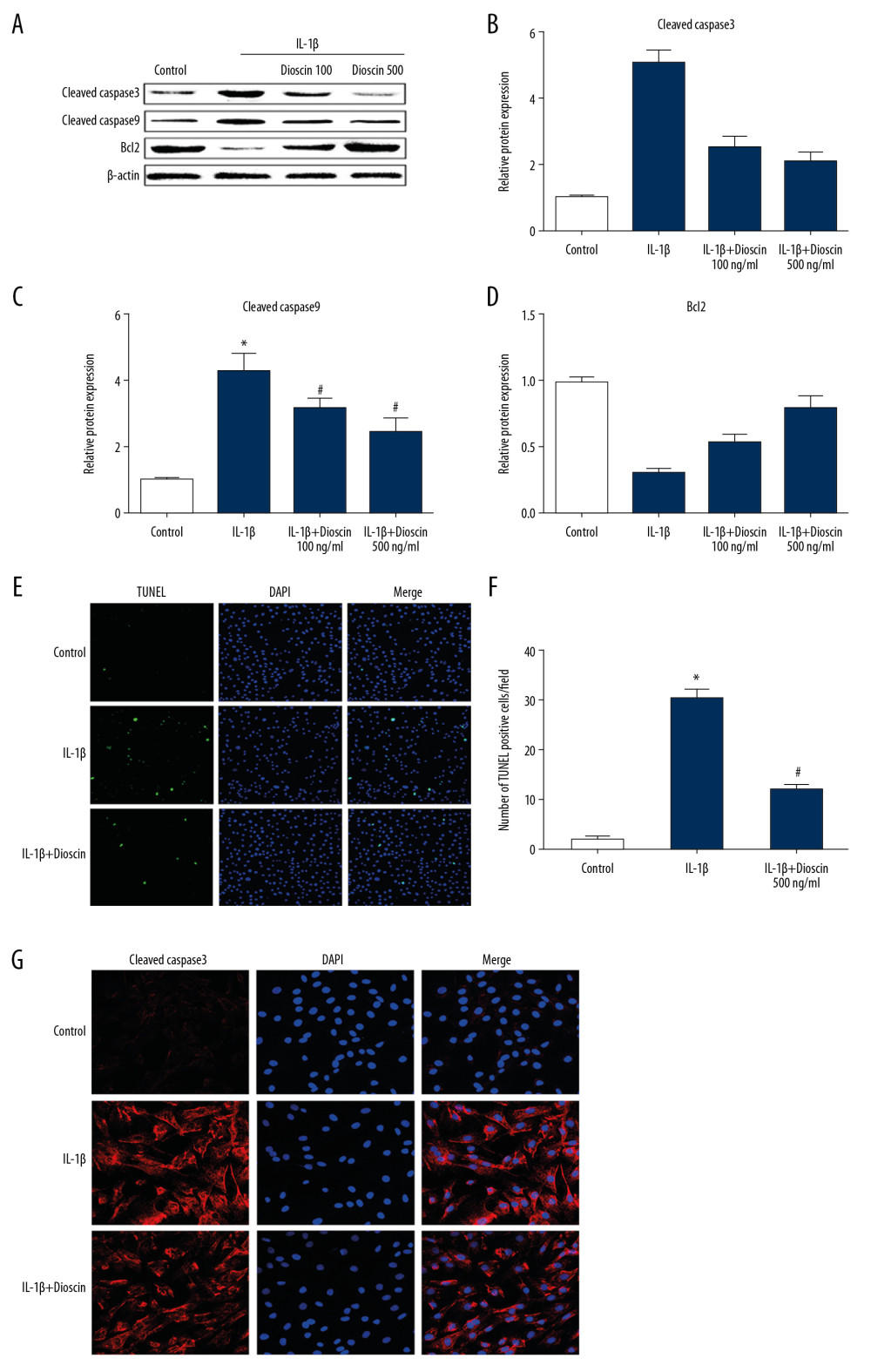 Figure 3. Effect of dioscin on IL-1β–activated apoptotic activity in human NP cells. (A, B) Apoptotic activity was evaluated using the TUNEL method. (C–F) Western blotting and quantitative analysis of cleaved caspase-3, -9, and Bcl-2 expressions. (G) Fluorescence immunostaining of cleaved caspase-3 (red) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.
Figure 3. Effect of dioscin on IL-1β–activated apoptotic activity in human NP cells. (A, B) Apoptotic activity was evaluated using the TUNEL method. (C–F) Western blotting and quantitative analysis of cleaved caspase-3, -9, and Bcl-2 expressions. (G) Fluorescence immunostaining of cleaved caspase-3 (red) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5. 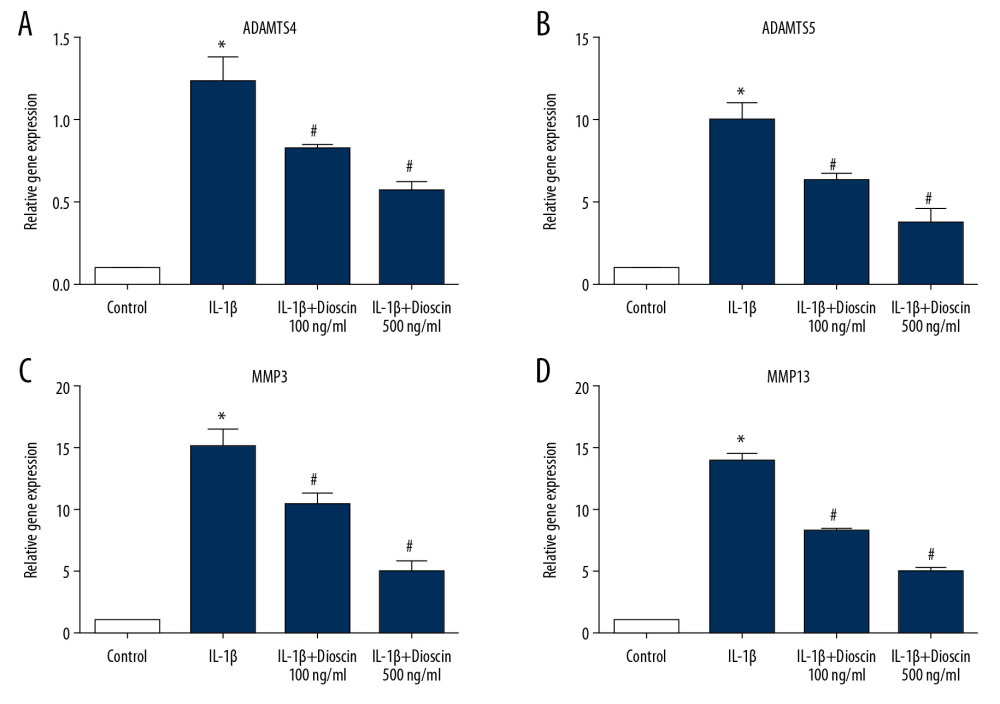 Figure 4. Dioscin suppressed IL-1β-activated catabolic activity in human NP cells. (A–D) Relative mRNA level of (A, B) ADAMTS4 and ADAMTS5, and (C, D) MMP3, MMP13 were measured by PCR. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.
Figure 4. Dioscin suppressed IL-1β-activated catabolic activity in human NP cells. (A–D) Relative mRNA level of (A, B) ADAMTS4 and ADAMTS5, and (C, D) MMP3, MMP13 were measured by PCR. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5. 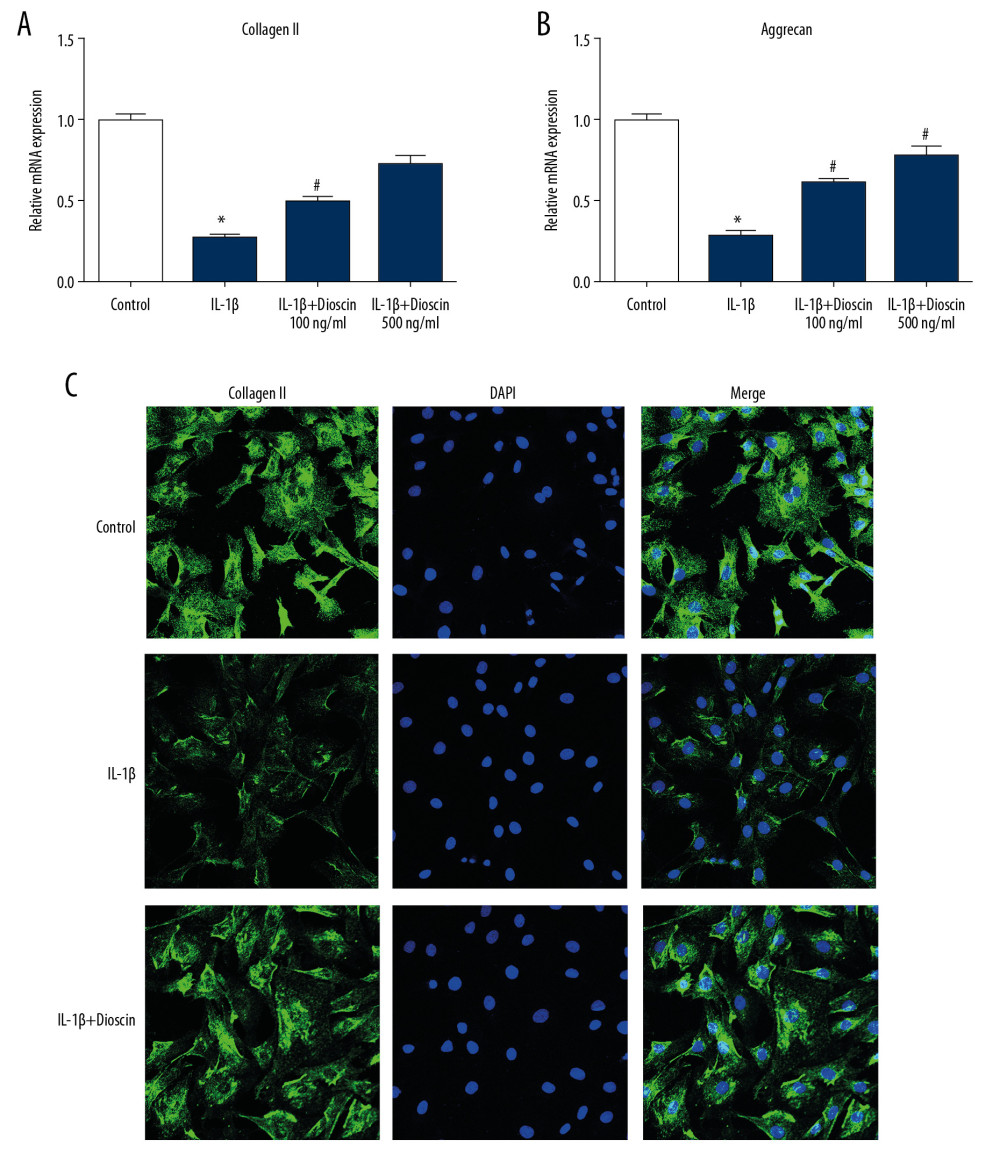 Figure 5. Dioscin attenuated ECM degradation in IL-1β-stimulated human NP cells. (A, B) Relative mRNA level of (A) collagen II and (B) aggrecan were examined by PCR. (C) Fluorescence immunostaining of collagen II protein (green) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.
Figure 5. Dioscin attenuated ECM degradation in IL-1β-stimulated human NP cells. (A, B) Relative mRNA level of (A) collagen II and (B) aggrecan were examined by PCR. (C) Fluorescence immunostaining of collagen II protein (green) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5. 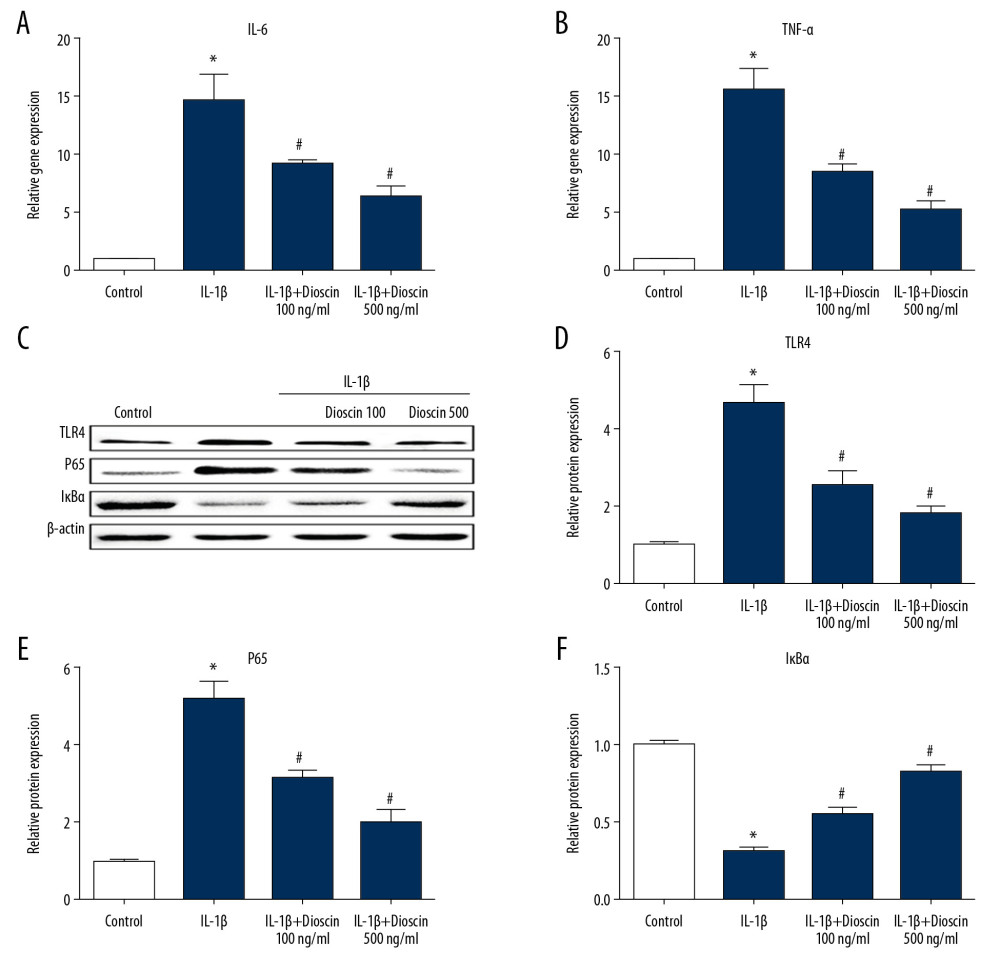 Figure 6. Dioscin suppressed IL-1β-induced release of proinflammatory factors via regulation of the NF-κB pathway in human NP cells. (A, B) Relative mRNA level of (A) IL-6 and (B) TNF-α were examined by PCR. (C–F) Western blotting and quantitative analysis of TLR4, p65, and IκBα. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.
Figure 6. Dioscin suppressed IL-1β-induced release of proinflammatory factors via regulation of the NF-κB pathway in human NP cells. (A, B) Relative mRNA level of (A) IL-6 and (B) TNF-α were examined by PCR. (C–F) Western blotting and quantitative analysis of TLR4, p65, and IκBα. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5. 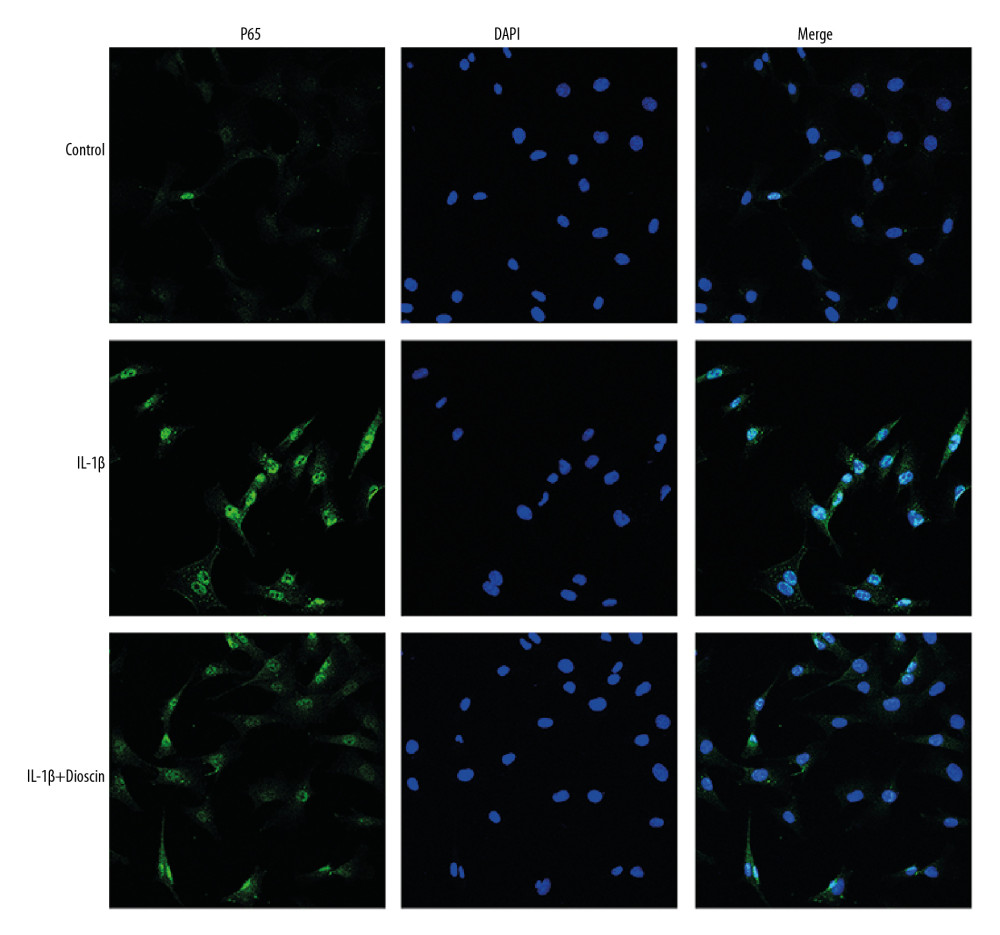 Figure 7. Dioscin inhibited nuclear translocation of NF-κB in human NP cells. Fluorescence immunostaining of p65 (green) and nucleus (blue).
Figure 7. Dioscin inhibited nuclear translocation of NF-κB in human NP cells. Fluorescence immunostaining of p65 (green) and nucleus (blue). 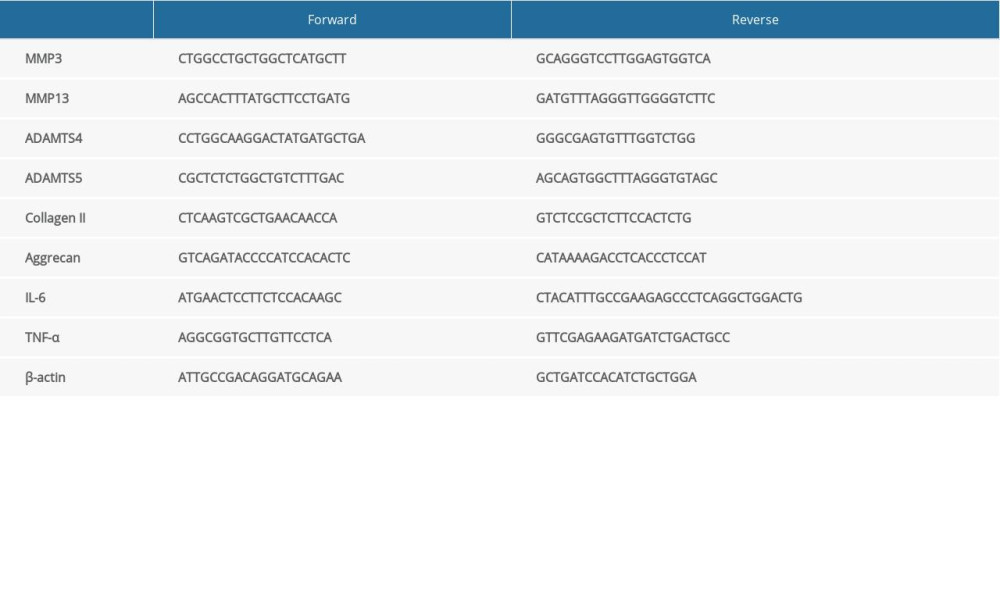
References
1. Sheng SR, Wang XY, Xu HZ, Anatomy of large animal spines and its comparison to the human spine: A systematic review: Eur Spine J, 2010; 19; 46-56
2. McBeth J, Jones K, Epidemiology of chronic musculoskeletal pain: Best Pract Res Clin Rheumatol, 2007; 21; 403-25
3. Freemont AJ, The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain: Rheumatology (Oxford), 2009; 48; 5-10
4. Jiang L, Zhang X, Zheng X, Apoptosis, senescence, and autophagy in rat nucleus pulposus cells: Implications for diabetic intervertebral disc degeneration: J Orthop Res, 2013; 31; 692-702
5. Zhou KL, Zhou YF, Wu K, Stimulation of autophagy promotes functional recovery in diabetic rats with spinal cord injury: Sci Rep, 2015; 5; 17130
6. Hayes AJ, Benjamin M, Ralphs JR, Extracellular matrix in development of the intervertebral disc: Matrix Biol, 2001; 20; 107-21
7. Hutton WC, Ganey TM, Elmer WA, Does long-term compressive loading on the intervertebral disc cause degeneration?: Spine (Phila Pa 1976), 2000; 25; 2993-3004
8. Sivan SS, Tsitron E, Wachtel E, Aggrecan turnover in human intervertebral disc as determined by the racemization of aspartic acid: J Biol Chem, 2006; 281; 13009-14
9. Weiler C, Nerlich AG, Bachmeier BE, Boos N, Expression and distribution of tumor necrosis factor alpha in human lumbar intervertebral discs: A study in surgical specimen and autopsy controls: Spine (Phila Pa 1976), 2005; 30; 44-53 discussion 54
10. Le Maitre CL, Freemont AJ, Hoyland JA, The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration: Arthritis Res Ther, 2005; 7; R732-45
11. Zu B, Pan H, Zhang XJ, Yin ZS, Serum levels of the inflammatory cytokines in patients with lumbar radicular pain due to disc herniation: Asian Spine J, 2016; 10; 843-49
12. Dinarello CA, Interleukin-1 in the pathogenesis and treatment of inflammatory diseases: Blood, 2011; 117; 3720-32
13. Wuertz K, Vo N, Kletsas D, Boos N, Inflammatory and catabolic signalling in intervertebral discs: The roles of NF-kappaB and MAP kinases: Eur Cell Mater, 2012; 23; 103-19 discussion 119–20
14. Brautbar N, Williams J, Industrial solvents and liver toxicity: Risk assessment, risk factors and mechanisms: Int J Hyg Environ Health, 2002; 205; 479-91
15. Qi M, Zheng L, Qi Y, Dioscin attenuates renal ischemia/reperfusion injury by inhibiting the TLR4/MyD88 signaling pathway via up-regulation of HSP70: Pharmacol Res, 2015; 100; 341-52
16. Zhang W, Yin L, Tao X, Dioscin alleviates dimethylnitrosamine-induced acute liver injury through regulating apoptosis, oxidative stress and inflammation: Environ Toxicol Pharmacol, 2016; 45; 193-201
17. Zhao L, Tao X, Qi Y, Protective effect of dioscin against doxorubicin-induced cardiotoxicity via adjusting microRNA-140-5p-mediated myocardial oxidative stress: Redox Biol, 2018; 16; 189-98
18. Qiao Y, Xu L, Tao X, Protective effects of dioscin against fructose-induced renal damage via adjusting Sirt3-mediated oxidative stress, fibrosis, lipid metabolism and inflammation: Toxicol Lett, 2018; 284; 37-45
19. Tao X, Yin L, Xu L, Peng J, Dioscin: A diverse acting natural compound with therapeutic potential in metabolic diseases, cancer, inflammation and infections: Pharmacol Res, 2018; 137; 259-69
20. Mao Z, Han X, Chen D, Potent effects of dioscin against hepatocellular carcinoma through regulating TP53-induced glycolysis and apoptosis regulator (TIGAR)-mediated apoptosis, autophagy, and DNA damage: Br J Pharmacol, 2019; 176; 919-37
21. Zheng L, Han X, Hu Y, Dioscin ameliorates intestinal ischemia/reperfusion injury via adjusting miR-351-5p/MAPK13-mediated inflammation and apoptosis: Pharmacol Res, 2019; 139; 431-39
22. Hu Y, Mao Z, Xu L, Protective effect of dioscin against intestinal ischemia/reperfusion injury via adjusting miR-351-5p-mediated oxidative stress: Pharmacol Res, 2018; 137; 56-63
23. Yao H, Tao X, Xu L, Dioscin alleviates non-alcoholic fatty liver disease through adjusting lipid metabolism via SIRT1/AMPK signaling pathway: Pharmacol Res, 2018; 131; 51-60
24. Zhao X, Yin L, Fang L, Protective effects of dioscin against systemic inflammatory response syndromevia adjusting TLR2/MyD88/NFkappab signal pathway: Int Immunopharmacol, 2018; 65; 458-69
25. Tao X, Sun X, Yin L, Dioscin ameliorates cerebral ischemia/reperfusion injury through the downregulation of TLR4 signaling via HMGB-1 inhibition: Free Radic Biol Med, 2015; 84; 103-15
26. Risbud MV, Guttapalli A, Stokes DG, Nucleus pulposus cells express HIF-1 alpha under normoxic culture conditions: A metabolic adaptation to the intervertebral disc microenvironment: J Cell Biochem, 2006; 98; 152-59
27. Deyo RA, Tsui-Wu YJ, Descriptive epidemiology of low-back pain and its related medical care in the United States: Spine (Phila Pa 1976), 1987; 12; 264-68
28. Brinjikji W, Diehn FE, Jarvik JG, MRI findings of disc degeneration are more prevalent in adults with low back pain than in asymptomatic controls: A systematic review and meta-analysis: Am J Neuroradiol, 2015; 36; 2394-99
29. Fernandez-Moure J, Moore CA, Kim K, Novel therapeutic strategies for degenerative disc disease: Review of cell biology and intervertebral disc cell therapy: SAGE Open Med, 2018; 6; 2050312118761674
30. Qin C, Lv Y, Zhao H, MicroRNA-149 suppresses inflammation in nucleus pulposus cells of intervertebral discs by regulating MyD88: Med Sci Monit, 2019; 25; 4892-900
31. Vergroesen PP, Kingma I, Emanuel KS, Mechanics and biology in intervertebral disc degeneration: A vicious circle: Osteoarthritis Cartilage, 2015; 23; 1057-70
32. Feng C, Yang M, Lan M, ROS: Crucial intermediators in the pathogenesis of intervertebral disc degeneration: Oxid Med Cell Longev, 2017; 2017 5601593
33. Kong L, Sun M, Jiang Z, MicroRNA-194 inhibits lipopolysaccharide-induced inflammatory response in nucleus pulposus cells of the intervertebral disc by targeting TNF receptor-associated factor 6 (TRAF6): Med Sci Monit, 2018; 24; 3056-67
34. Lv J, Li S, Wan T, Inhibition of microRNA-30d attenuates the apoptosis and extracellular matrix degradation of degenerative human nucleus pulposus cells by up-regulating SOX9: Chem Biol Interact, 2018; 296; 89-97
35. Kim HA, Blanco FJ, Cell death and apoptosis in ostearthritic cartilage: Current Drug Targets, 2007; 8; 333-45
36. Qin J, Kang Y, Xu Z, Dioscin prevents the mitochondrial apoptosis and attenuates oxidative stress in cardiac H9c2 cells: Drug Res (Stuttg), 2014; 64; 47-52
37. Wang J, Chen H, Cao P, Inflammatory cytokines induce caveolin-1/beta-catenin signalling in rat nucleus pulposus cell apoptosis through the p38 MAPK pathway: Cell Prolif, 2016; 49; 362-72
38. Lv F, Huang Y, Lv W, MicroRNA-146a ameliorates inflammation via TRAF6/NF-kappaB pathway in intervertebral disc cells: Med Sci Monit, 2017; 23; 659-64
39. Shah BS, Burt KG, Jacobsen T, High mobility group box-1 induces pro-inflammatory signaling in human nucleus pulposus cells via toll-like receptor 4-dependent pathway: J Orthop Res, 2018; 37; 220-31
40. Gomez R, Villalvilla A, Largo R, TLR4 signalling in osteoarthritis – finding targets for candidate DMOADs: Nat Rev Rheumatol, 2015; 11; 159-70
41. Rajan NE, Bloom O, Maidhof R, Toll-like receptor 4 (TLR4) expression and stimulation in a model of intervertebral disc inflammation and degeneration: Spine (Phila Pa 1976), 2013; 38; 1343-51
42. Zhang L, Chen Q, Wang H, Andrographolide mitigates IL1betainduced human nucleus pulposus cells degeneration through the TLR4/MyD88/NFkappaB signaling pathway: Mol Med Rep, 2018; 18; 5427-36
43. Lorenz W, Buhrmann C, Mobasheri A, Bacterial lipopolysaccharides form procollagen-endotoxin complexes that trigger cartilage inflammation and degeneration: implications for the development of rheumatoid arthritis: Arthritis Res Ther, 2013; 15; R111
44. Bobacz K, Sunk IG, Hofstaetter JG, Toll-like receptors and chondrocytes: The lipopolysaccharide-induced decrease in cartilage matrix synthesis is dependent on the presence of toll-like receptor 4 and antagonized by bone morphogenetic protein 7: Arthritis Rheum, 2007; 56; 1880-93
45. Kim JS, Ellman MB, Yan D, Lactoferricin mediates anti-inflammatory and anti-catabolic effects via inhibition of IL-1 and LPS activity in the intervertebral disc: J Cell Physiol, 2013; 228; 1884-96
46. Yang W, Yu XH, Wang C, Interleukin-1beta in intervertebral disk degeneration: Clin Chim Acta, 2015; 450; 262-72
47. Kang JD, Stefanovic-Racic M, McIntyre LA, Toward a biochemical understanding of human intervertebral disc degeneration and herniation. Contributions of nitric oxide, interleukins, prostaglandin E2, and matrix metalloproteinases: Spine (Phila Pa 1976), 1997; 22; 1065-73
48. Wu S, Xu H, Peng J, Potent anti-inflammatory effect of dioscin mediated by suppression of TNF-alpha-induced VCAM-1, ICAM-1and EL expression via the NF-kappaB pathway: Biochimie, 2015; 110; 62-72
Figures
Figure 1. The molecular structure of dioscin.Figure 2. Effects of dioscin on IL-1β-activated cell viability in human NP cells. (A) The cytotoxic effects of dioscin on human NP cells were examined at different concentrations by the CCK-8 method. (B) CCK-8 analysis of dioscin-treated human NP cells stimulated by IL-1β. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.Figure 3. Effect of dioscin on IL-1β–activated apoptotic activity in human NP cells. (A, B) Apoptotic activity was evaluated using the TUNEL method. (C–F) Western blotting and quantitative analysis of cleaved caspase-3, -9, and Bcl-2 expressions. (G) Fluorescence immunostaining of cleaved caspase-3 (red) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.Figure 4. Dioscin suppressed IL-1β-activated catabolic activity in human NP cells. (A–D) Relative mRNA level of (A, B) ADAMTS4 and ADAMTS5, and (C, D) MMP3, MMP13 were measured by PCR. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.Figure 5. Dioscin attenuated ECM degradation in IL-1β-stimulated human NP cells. (A, B) Relative mRNA level of (A) collagen II and (B) aggrecan were examined by PCR. (C) Fluorescence immunostaining of collagen II protein (green) and nucleus (blue). The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.Figure 6. Dioscin suppressed IL-1β-induced release of proinflammatory factors via regulation of the NF-κB pathway in human NP cells. (A, B) Relative mRNA level of (A) IL-6 and (B) TNF-α were examined by PCR. (C–F) Western blotting and quantitative analysis of TLR4, p65, and IκBα. The results are presented as the means ±SD. * P<0.05 relative to the control group; # P<0.05 relative to the IL-1β group, n=5.Figure 7. Dioscin inhibited nuclear translocation of NF-κB in human NP cells. Fluorescence immunostaining of p65 (green) and nucleus (blue). In Press
Clinical Research
Body Weight and Insulin Resistance Indicators Among ChildrenMed Sci Monit In Press; DOI: 10.12659/MSM.951434
Clinical Research
Comparison of Radiographic Cervical Sagittal Alignment Parameters in Patients With Nonspecific Neck Pain, D...Med Sci Monit In Press; DOI: 10.12659/MSM.952950
Clinical Research
Combined Fibrinogen and Urinary α1-Microglobulin as Predictors of Respiratory Tract Infection in Children w...Med Sci Monit In Press; DOI: 10.12659/MSM.951066
Database Analysis
Evaluation of Salivary Total Oxidant Status (TOS) and Total Antioxidant Status (TAS) in Orthodontic Patient...Med Sci Monit In Press; DOI: 10.12659/MSM.952052
Most Viewed Current Articles
17 Jan 2024 : Review article 14,175,576
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,756,620
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,465,966
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,651
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387