16 February 2021: Animal Study
Effects of Diabetes Mellitus on Sperm Quality in the Db/Db Mouse Model and the Role of the FoxO1 Pathway
Jisheng Wang12ABDE, Binghao Bao12ABE, Junlong Feng12ABCE, Qi Zhao12BCF, Hengheng Dai12BDF, Fanchao Meng12BF, Sheng Deng12DF, Bin Wang2AG*, Haisong Li2AEDOI: 10.12659/MSM.928232
Med Sci Monit 2021; 27:e928232






Abstract
BACKGROUND: Studies have shown that diabetes mellitus (DM) has a negative impact on male reproductive function, which may lead to changes in the testis and epididymis and a decline in semen quality.
MATERIAL AND METHODS: We performed animal experiments with 6 diabetic db/db mice as the model group (group B) and 6 C57BL/6J mice as the control group (group A). After adaptive feeding for 7 days, the sperm quality of each group was measured. Concurrently, the morphology of the mouse testis was observed by hematoxylin-eosin (H&E) staining. The expression of the PI3K, Akt, FoxO1, FasL, IL-6, and Stat3 proteins and mRNAs in the testicular tissue was detected by western blotting and RT-qPCR.
RESULTS: The number of spermatozoa and sperm motility of group A was significantly higher than that of group B (P<0.05). H&E staining of the testicular tissue showed the seminiferous tubules in group B mice were damaged to varying degrees and the seminiferous tubules were sparsely arranged. Compared with those of group A, the expression levels of PI3K, Akt, and Stat3 proteins and mRNAs in group B were significantly lower (P<0.05), while the expression levels of FoxO1, FasL, and IL-6 proteins and mRNAs in group B mice were significantly higher (P<0.05).
CONCLUSIONS: This study demonstrated that DM inhibited the expression of PI3K, Akt, and Stat3 proteins and mRNAs in the FoxO1 pathway and promoted the expression of FoxO1, FasL, and IL-6 proteins and mRNAs, leading to abnormal apoptosis of testicular tissue cells and functional damage, and eventually spermatogenic dysfunction.
Keywords: Diabetes Mellitus, Infertility, Diabetes Complications, Diabetes Mellitus, Experimental, Forkhead Box Protein O1, semen analysis, Sperm Count, Sperm Motility, Spermatogenesis, Spermatozoa, Testis
Background
According to the World Health Organization (WHO), infertility is defined as the inability to conceive after 1 year of unprotected intercourse [1]. Epidemiological investigation showed that the prevalence of infertility is approximately 10% to 12%, with more than 40% of infertility attributable to male factors [2]. Approximately 65% to 70% of the etiology and pathogenesis of male infertility are still unknown, and most patients show only a decline in sperm quality [3]. Some cases of male infertility may be induced by other secretory diseases, with symptoms including decreased sperm motility and quantity. Diabetes mellitus (DM) is recognized as one of the 3 major chronic diseases by the WHO. About 415 million people worldwide have diabetes or are at risk of developing early diabetes [4]. Data show that the prevalence of diabetes in China is as high as 10.9% and the number of patients with diabetes in China is the highest in the world [5]. Studies have shown that DM has a negative impact on male reproductive function, which may lead to changes in the testis and epididymis and a decline in semen quality, such as changes in semen volume, sperm count, sperm motility, and sperm morphology [6,7]. However, the exact mechanism of DM-induced decline in male semen quality is not fully understood. Studies suggest that hyperglycemia may have adverse effects on male reproductive function by affecting the endocrine hormones in the hypothalamic-pituitary-testicular axis and prostate development [8].
The FoxO1 signaling pathway is the main pathway for insulin signal transduction and regulating blood glucose [9]. FoxO1 is one of the main members of the subfamily of FoxO, which is widespread in mammals and is a direct downstream signaling molecule of the PI3K/Akt signaling pathway. The PI3K/Akt signaling pathway is involved in the regulation of a variety of biological processes, including the cell cycle, cell growth, cell proliferation, and cell metabolism [10]. The anti-apoptotic effect of the PI3K/Akt signaling pathway may be related to the regulation of downstream protein FoxO expression. When PI3K/Akt is activated, Akt enters the nucleus to phosphorylate FoxO1, and the 14-3-3 binding protein binds to phosphorylated FoxO1 and translocates out of the nucleus. FoxO1 loses its transcriptional activity, which in turn increases cell proliferation and anti-apoptotic ability [11]. FasL has been considered to be a key regulator of testicular germ-cell apoptosis, which plays an important regulatory role in testicular spermatogenic function [12]. Interleukin 6 (IL-6) is a multifunctional cytokine involved in both proinflammatory and antiinflammatory actions, and studies have found that the concentration of IL-6 in the seminal plasma of men with infertility is significantly higher than that of men with normal fertility [13]. Signal transducer and activator of transcription (STAT)3 is widely expressed in the central nervous system during development and adulthood, and mice with knockout of STAT3 were found to have hypogonadism and testicular and seminal vesicle atrophy [14].
Network pharmacology is based on the rapid development of systems biology and computer technology. Recently, network pharmacology has become a powerful tool combined with pharmacology. Through bioinformatic predictions, we found that the FoxO1 signaling pathway may be a potential mechanism for DM to affect spermatogenesis. Therefore, based on the prediction results of bioinformatics technology combined with animal experiments, the aim of this study was to determine the mechanism of DM-causing oligospermia. The overview of our research process is shown in Figure 1.
Material and Methods
IDENTIFICATION OF MOLECULAR TARGETS:
We identified the molecular targets of diabetes and male infertility using the GeneCards database (
PROTEIN–PROTEIN INTERACTION NETWORK ANALYSIS:
The STRING database (
NETWORK CONSTRUCTION AND ANALYSIS:
Cytoscape software (version 3.7.1,
GENE ONTOLOGY AND KYOTO ENCYCLOPEDIA OF GENES AND GENOMES ENRICHMENT ANALYSIS:
The biological information annotation database (DAVID,
ETHICAL APPROVAL OF THE STUDY PROTOCOL:
All experimental protocols conformed to the guidelines approved by the Animal Ethics Committee of Dongzhimen Hospital, Beijing University of Traditional Chinese Medicine (approval nos. 17–27).
ANIMAL EXPERIMENTS:
Six 8-week-old db/db male mice were selected as the diabetic group (group B), and 6 C57BL/6J male mice of the same age were selected as the normal control group (group A), with an average weight of 25 g to 40 g. All mice were purchased by our laboratory [animal license no.: SCXK (Beijing) 2016-0010]. The mice were adaptively fed for 7 days before the experiments. Feeding and experiments were conducted in the animal laboratory of the Animal Center of Dongzhimen Hospital, Beijing University of Traditional Chinese Medicine.
REAGENTS AND EQUIPMENT:
The following were used in the experiments:
SDS polyacrylamide gel (SDS)-PAGE gel kit (GenePool, GPP1816, Beijing, China); SDS-PAGE loading buffer (5×) (GenePool); protein extraction kit (GenePool); tris-glycine running buffer (5×) (GenePool); total RNA extraction kit (DNase I) (GenePool, Cat no. GPQ1801); mRNA cDNA synthesis kit (GenePool, Cat no. GPQ1803, Beijing, China); mRNA/lncRNA qPCR kit (GenePool, Cat no. GPQ1808); RNA loading buffer (5×) (GenePool, Cat no. GPQ1813). PI3K antibody (Abcam, ab191606; 1: 1000 dilution, Cambridge, UK); p-Akt antibody (Bioss, 0876R, 1: 2000 dilution, Woburn, Massachusetts, USA); P-FoxO1 antibody (Bioss,20095R, 1: 2000 dilution); FasL antibody (Bioss, bs-0216R; 1: 500 dilution); p-Stat3 antibody (Bioss, 22386R, 1: 2000 dilution); IL-6 antibody (Abcam, ab208113; 1: 500 dilution); Actin antibody (Abcam, ab6276; 1: 3000 dilution); goat anti-mouse IgG, HRP (Abcam, ab6789; 1: 5000 dilution); goat anti-rabbit IgG, HRP (Abcam, ab6721; 1: 5000 dilution); electrophoresis apparatus (CAVOY, PP-1150); spectrophotometer (NANODROP 2000, Thermo Scientific); real time PCR (Line Gene 9600 Plus, Bioer Technology); and double vertical electrophoresis cell (CAVOY, MP-8001).
CONFIRMATION OF MOUSE MODEL:
The mice in both groups were fed a normal diet for 7 days, testing for blood glucose was done by tail vein cutting, blood glucose was measured by a Roche blood glucose meter and blood glucose test paper (Roche, Germany), and model validation was performed by a general observation index and blood glucose level. Polydipsia, polyphagia, polyuria, and random blood glucose >16.7 mmol/L confirmed the DM mice model.
PREPARATION OF BLOOD AND TISSUE SAMPLES:
At the end of the experiment, mice were weighed and anesthetized using sodium pentobarbital (50 mg/kg via intraperitoneal injection). Blood was drawn from the tail vein to measure blood glucose levels, and testicular and epididymal tissues were removed. The epididymis was used to measure sperm motility and sperm count. Western blotting and qRT-PCR were performed on the left testis, and the right testis was used to observe tissue morphology.
DETERMINATION OF SPERM QUALITY:
The epididymal tissues were excised to remove excess fat, and the WLJY-9000 Weili sperm quality detection system (Xingrong Technology Co., Ltd., Beijing, China) was used for testing. The system was debugged and the temperature was set to 37 °C, according to the manufacturer’s instructions. Then, for semen specimen testing, the counting plate was placed on the thermostat for heating. After semen liquefaction, semen analysis specimens were prepared, and 5 μL of the quantified semen was dripped onto the semen pool of the clean counting plate with a micropipette and then covered with a cover glass. The microscope was mounted on a thermostatic plate on the stage and analyzed. Specific observation methods were conducted according to WHO criteria: 10 fields of view were randomly selected and the numbers of sperm in 10 large squares were counted. The average value was 106 per mL.
TESTICULAR MORPHOLOGY:
After the testicular tissue was washed, a portion of the testis was embedded in paraffin, cut into thin slices (5 μm thick), sectioned with hematoxylin-eosin (H&E) staining, and observed and photographed under a light microscope equipped with a digital tube (Olympus BX51TF, Tokyo, Japan).
WESTERN BLOTTING DETECTION:
To detect the expression levels of PI3K, Akt, FoxO1, FasL, Stat3, and IL-6 protein, we removed a portion of testicular tissue from each mouse. These tissues were placed into ice-cold tissue lysis buffer and homogenized. The homogenate was then centrifuged at 12 000 rpm for 10 min to allow collection of the supernatant. Protein concentration in each lysate was determined using a bicinchoninic acid (BCA) protein assay kit and the concentrations were adjusted with radio-immunoprecipitation assay buffer and boiled for 5 min. Each sample was then separated by 10% SDS-PAGE electrophoresis and transferred to a polyvinylidene fluoride (PVDF) membrane. The primary antibody was diluted, incubated at 4°C overnight, and washed with Tris-buffered saline with Tween (TBST) at room temperature on a decolorizing shaker 3 times for 5 min each time. Next, the secondary antibody was diluted 3000 times with TBST. After incubating at room temperature for 30 min, it was washed 3 times with TBST on a decolorizing shaker at room temperature for 5 min each time. The reagents ECLA and ECLB were mixed in a medium volume in a centrifuge tube, and after fully contacting the PVDF membrane, they were placed in an automated chemiluminescence imaging system to read the data [15].
RT-QPCR DETECTION:
A PCR instrument (SLAN-96P, SJ, CN) was used for qPCR. When extracting total RNA, TRIzol Reagent (15596-026; Invitrogen Life Technologies, Carlsbad, CA, USA) was added to the homogenized tissue to maintain the integrity of the RNA in the sample and to inhibit RNA degradation. The purity and concentration of total RNA was determined, and cDNA was prepared by reverse transcription. Primer sequences are shown in Table 1. Samples were added to 96-well plates and amplified by fluorescence quantitative PCR. The number of cycles (Ct value) experienced by the fluorescent signal when the set threshold was reached in each reaction tube was recorded The difference in the expression of the gene was determined by computing the multiple of the target gene relative to the reference, with the relative quantification (RQ) method (RQ=2–ΔΔCt).
STATISTICAL ANALYSIS:
SPSS version 20.0 (SPSS Inc, Chicago, Illinois, USA) was used for all statistical analyses. Data were expressed as mean±SD. If the data were normally distributed, a
Results
COLLECTION OF TARGETS:
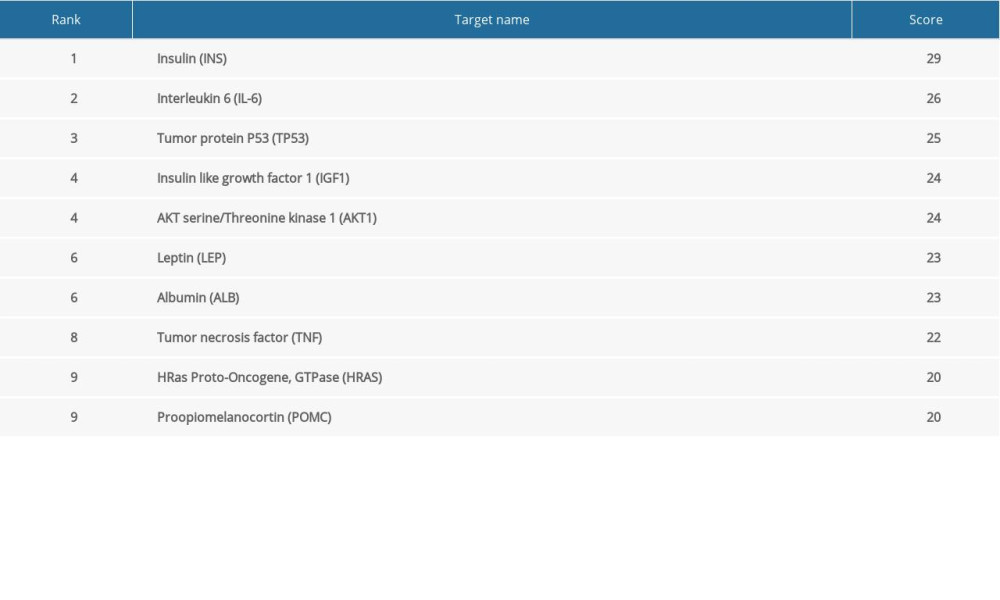
The flowchart of our study is shown in Figure 1. Genecard screening allowed us to identify 300 diabetes targets and 300 male infertility targets. Following intersection, 37 targets related to diabetes and male infertility were identified (Figure 2A).
CONSTRUCTION AND TOPOLOGICAL ANALYSIS OF THE DIABETES-MALE INFERTILITY NETWORK AND THE PPI NETWORK:
Cytoscape software was used to construct a diabetes-male infertility network (Figure 2B). The 37 overlapping targets for diabetes and male infertility were then analyzed using the STRING database. Cytoscape software was used to construct a PPI network for the 37 targets (Figure 2C). The ‘cytohubba’ plug-in was used to analyze the PPI network and identify the top 10 key targets (Figure 2D, Table 2).
GENE ONTOLOGY BIOLOGICAL PROCESS ENRICHMENT ANALYSIS:
Enrichment analysis of the 37 targets using DAVID version 6.8 further indicated the involvement of 76 cell biological processes, 9 cell components, 17 molecular functions, and 66 signaling pathways. The top 10 biological functions and signaling pathways were then selected based on their P values (Figure 3).
DIABETIC MOUSE MODEL:
Through 3 consecutive random measurements of the tail-vein blood glucose in mice, we found that the 6 db/db mice (group B) had blood glucose levels higher than 16.7 mmol/L, and the 6 C57BL/6J mice (group A) had blood glucose levels lower than 16.7 mmol/L (Figure 4A, 4B). At the same time, the db/db mice showed polydipsia, polyphagia, and polyuria, which are characteristic manifestations of DM.
SPERM QUALITY:
The number of sperm in group A was significantly higher than that in group B (P<0.05) (Figure 4C, 4D). The sperm motility in group A was significantly higher than that in group B (P<0.05) (Figure 4E, 4F).The results showed that the semen quality of the diabetic mice was significantly lower than that of the control group.
TESTICULAR MORPHOLOGY:
The seminiferous tubules of group A mice were closely arranged, and the boundary between the basement membrane and stroma was clear and smooth. Sertoli cells and spermatozoa in the seminiferous tubules were arranged neatly and abundantly. The seminiferous tubules of group B mice were injured to different degrees, and the seminiferous tubules were sparsely arranged, showing interstitial edema, and their epitheliums were separated from the matrix. The number of Sertoli cells in the testes was significantly lower and arranged loosely in group B mice, and the number of spermatozoa was significantly less, even occasional spermatozoa (Figure 5).
EXPRESSION OF PI3K, AKT, FOXO1, FASL, STAT3, AND IL-6 PROTEINS IN MOUSE TESTICULAR TISSUE:
Western blotting was used to detect the expression of related proteins in testicular tissues of mice in both groups. By analyzing the gray value of expression, we found that the PI3K protein level of mice in group A was significantly lower than that of group B (P<0.05). Compared with group A, the Akt protein expression of mice in group B was significantly lower (P<0.05). Mice in group B had significantly lower Akt protein expression than group A (P<0.05), and FoxO1 protein expression was significantly higher (P<0.05). FasL and IL-6 protein expression was significantly higher in group B compared with that of group A (P<0.05); Stat3 protein expression was significantly lower in group B compared with that of group A (P<0.05) (Figure 6A–6H). The results showed that the FoxO1 pathway-related proteins were significantly changed in the testicular tissue of DM mice.
MRNA EXPRESSION OF PI3K, AKT, FOXO1, FASL, STAT3, AND IL-6 IN MOUSE TESTICULAR TISSUE:
Using RT-qPCR, we analyzed the expression values and found that, compared with that of group A, PI3K mRNA and Akt mRNA expression levels in group B mice was significantly lower (P<0.01). Compared with that of group A, FoxO1 mRNA and FasL expression in group B mice was significantly higher (P<0.01). The expression of mRNA was significantly higher (P<0.01) and the expression of Stat3 mRNA was significantly lower in group B compared with that of group A (P<0.01). The expression of IL-6 mRNA was significantly higher in group B compared with that of group A (P<0.01) (Figure 7A–7G).The results showed that the expression of FoxO1 pathway-related mRNAs in testicular tissue of DM mice was significantly higher than in the control mice.
Discussion
The reason for the sperm quality decline caused by DM is not clear. In this study, db/db mice, which were congenital type 2 diabetic mice due to the defect of the leptin receptor gene, were used as the model group. This is widely recognized as the preferred animal model to study human type 2 diabetes [16]. Bioinformatics analysis predicted that DM might cause sperm quality decline by inhibiting FoxO1 signaling pathway-related proteins and mRNAs. The results of our experimental validation are basically consistent with the predicted results of biological information.
The FoxO1 signaling pathway is the main pathway of insulin signal transduction and blood glucose regulation [17]. While the apoptosis of testicular germ cells is controlled by a variety of genes, and apoptosis can occur due to local or exogenous factors at each stage of spermatogenesis [18], it has been suggested that FoxO1 can induce apoptosis by inducing the expression of downstream apoptotic factor FasL, which binds specifically to Fas on the spermatogenic cell membrane and activates the cascade of related caspases through the death domain of intracellular peptide segments [19]. In addition, the overexpression of FasL can cause abnormal apoptosis of testicular tissue cells, which ultimately adversely affects the quality and quantity of sperm [20]. Our results showed that the expression of FoxO1, FasL protein and mRNA in testicular tissue of DM mice was significantly higher than that of the control mice, suggesting that the elevation of FoxO1, FasL protein and mRNA caused the decline of spermatogenic function in DM mice.
FoxO1 is simultaneously regulated by transcription factors from multiple upstream pathways, including the phosphorylation cascade regulation of PI3K/Akt and IL-6/Stat3 [21–22].
The PI3K-Akt signaling pathway can promote glucose uptake by peripheral target tissues and plays an important role in insulin secretion by islet beta cells [23]. PI3K acts on Akt downstream to phosphorylate it. Activation of Akt can lead to nuclear exclusion by phosphorylating Ser256 and Thr24 of FoxO1, thus inhibiting the transcriptional activity of FoxO1 and insulin resistance, increasing the body’s sensitivity to insulin and reducing blood glucose levels [24,25]. However, when the PI3K/Akt pathway is inhibited, Akt activity decreases, inhibits the phosphorylation of FoxO1 in the nucleus, and increases FoxO1 expression in the nucleus. Inhibition of the PI3K/Akt pathway not only reduces the sensitivity of the body to insulin, but also enhances the pro-apoptotic effect of FoxO1, induces the expression of downstream apoptotic factor FasL, and causes abnormal apoptosis of testicular tissue cells [26,27]. Our results showed that FoxO1 and FasL were regulated by upstream PI3K and Akt protein and mRNA, which destroyed the spermatogenic function of DM mice.
IL-6/Stat3, as upstream regulatory proteins of the FoxO1 pathway, also play an important role in FoxO1 pathway expression. Functional metabolism disorders in organisms under hyperglycemic conditions cause ischemia and hypoxia of local tissue and stimulate cells to release cytokines such as IL-6 in large quantities. These cytokines participate in the body’s inflammatory immune response, and their elevated levels will further aggravate the impairment of islet function in diabetic patients and exacerbate the course of diabetes. Binding of IL-6 to specific receptor complexes induces Stat3 phosphorylation and promotes FoxO1 expression, further leading to increased FasL expression [28,29]
To the best of our knowledge, the present study is the first experimental study to report the relationship between DM and oligospermia. At the same time, based on bioinformatics and experimental research, the method undertaken was scientific. This study clarifies the potential mechanism of DM-induced oligoasthenospermia in db/db male mice and provides a reliable basis for future experimental and clinical research. There were some limitations of this study. We did not detect the levels of serum sex hormones. Sex hormones have an important impact on spermatogenesis. However, this study was based on bioinformatics analysis, and there were no sex hormone-related indicators in the bioinformatics prediction results. We hope that scholars will investigate the changes in serum sex hormone levels in oligoasthenospermia caused by DM in a follow-up study. Meanwhile, based on the biological network module, 10 related pathways were predicted. If we had aimed to detect all pathways, more work would have been involved and a larger budget would have been required. Therefore, for experimental verification, we selected the FoxO1 pathway, which relevant literature suggests is the mechanism closest to that of the 2 diseases. Also, the scheme of having 6 mice in each of the 2 groups may have been a small sample size. However, this experiment is the first to verify the effect of type 2 diabetes on spermatogenesis in mice, which is still in the exploratory stage. In the future, we will do in-depth research by increasing the sample size and conducting drug intervention experiments.
Conclusions
The decrease of sperm quality may be related to the FoxO1 signaling pathway. DM can inhibit the expression of PI3K, Akt, and Stat3 proteins and mRNAs in the FoxO1 pathway and promote the expression of FoxO1, FasL, and IL-6 protein and mRNA, thus leading to abnormal apoptosis of testicular tissue cells and function damage and ultimately leading to spermatogenesis dysfunction.
Figures
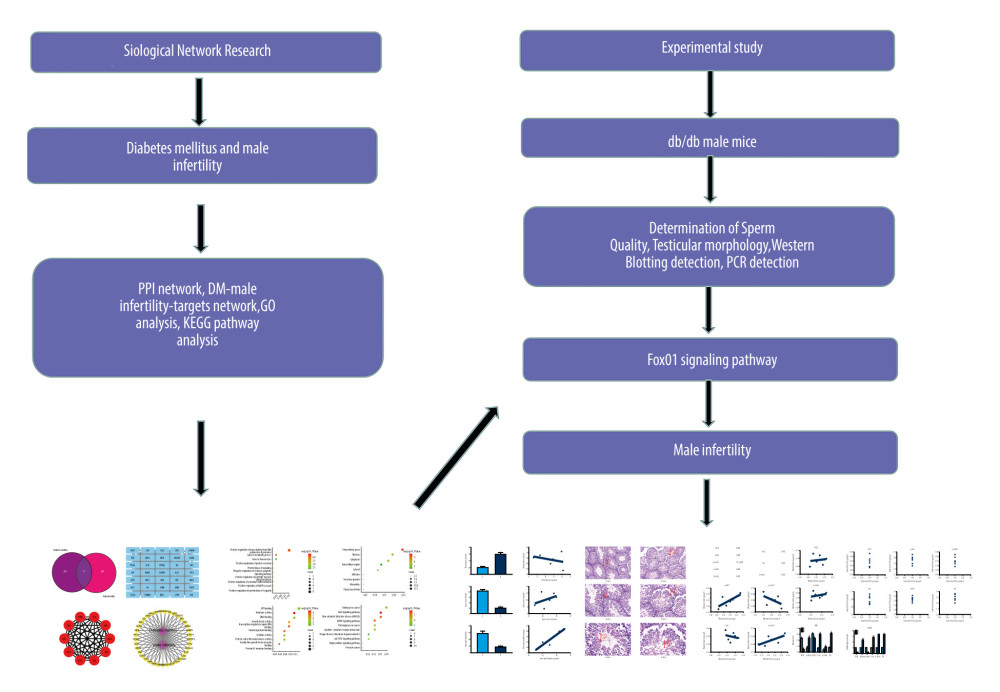 Figure 1. Overall process based on biological network research and animal experiments.
Figure 1. Overall process based on biological network research and animal experiments. 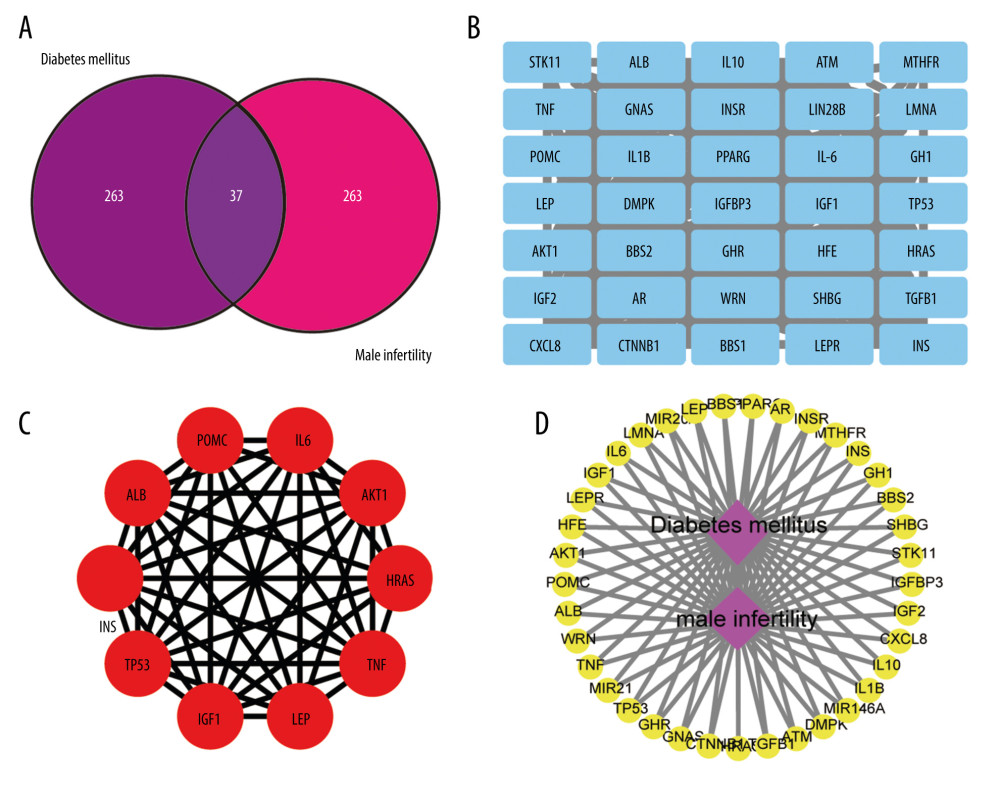 Figure 2. (A) Intersection of targets of diabetes mellitus and male infertility; (B) Diabetes mellitus-male infertility targets network built by Cytoscape (3.7.1); (C) protein–protein interaction (PPI) network built by Cytoscape (3.7.1); (D) PPI network processed by Cytoscape (3.7.1) plug-in (cytohubba).
Figure 2. (A) Intersection of targets of diabetes mellitus and male infertility; (B) Diabetes mellitus-male infertility targets network built by Cytoscape (3.7.1); (C) protein–protein interaction (PPI) network built by Cytoscape (3.7.1); (D) PPI network processed by Cytoscape (3.7.1) plug-in (cytohubba). 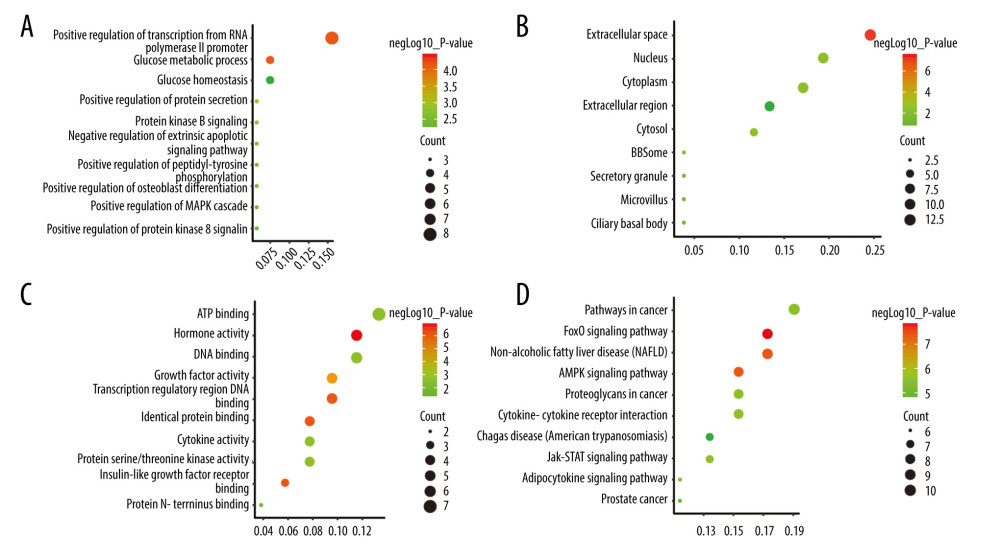 Figure 3. (A–D) Analyses of pathway enrichment using Gene Ontology and Kyoto Encyclopedia of Genes and Genomes databases. The Y axis represents the gene name and the X axis represents the richness factor. The size of the node is proportional to the number of genes. The node color is proportional to the P value.
Figure 3. (A–D) Analyses of pathway enrichment using Gene Ontology and Kyoto Encyclopedia of Genes and Genomes databases. The Y axis represents the gene name and the X axis represents the richness factor. The size of the node is proportional to the number of genes. The node color is proportional to the P value. 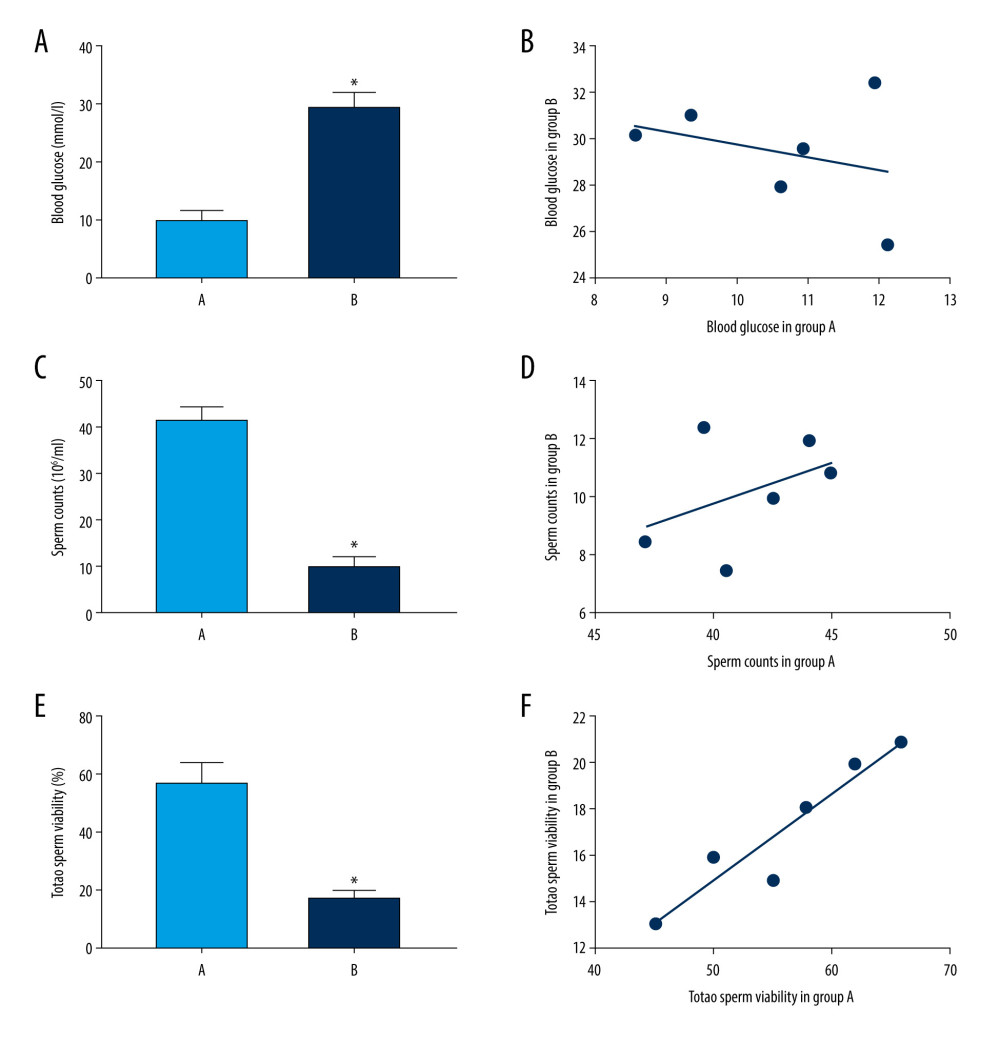 Figure 4. (A) Blood glucose of mice from groups A (control) and B (diabetic). (C, E) Sperm counts and total sperm viability in the 2 groups. (B, D, F) Scatter plot of blood glucose, sperm count and total sperm viability in the 2 groups of mice. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A, * P<0.05.
Figure 4. (A) Blood glucose of mice from groups A (control) and B (diabetic). (C, E) Sperm counts and total sperm viability in the 2 groups. (B, D, F) Scatter plot of blood glucose, sperm count and total sperm viability in the 2 groups of mice. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A, * P<0.05. 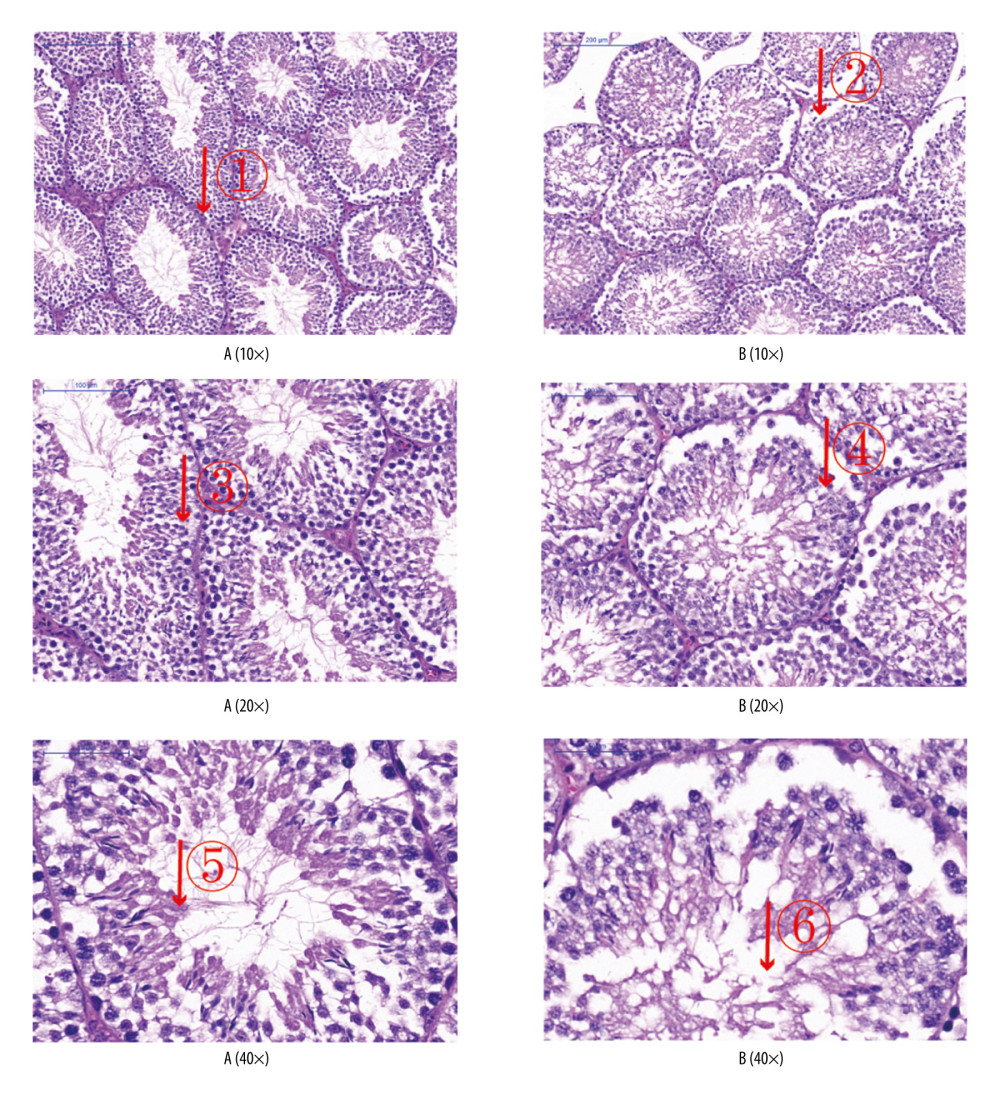 Figure 5. Analyses of testicular tissue in mice using hematoxylin-eosin (H&E) staining. The testicular tissue of mice was stained with H&E to observe pathologic changes in the testes under electron microscopy (n=6 animals per group). In group A: (i) The convoluted tubules are closely arranged, and the boundary between the basal membrane and the stroma is clear and even (arrow ?); (ii) all levels of spermatogenic cells and spermatogenic cells are arranged in order (arrows ?, ?). In group B: (i) Spermatogenic tubules are injured to different degrees, and the seminiferous tubules were sparsely arranged, showing interstitial edema, and their epithelium was separated from the matrix (arrow ?); (ii) the number of Sertoli cells in testes was significantly reduced and arranged loosely. The number of spermatozoa was significantly reduced (arrows ?, ?).
Figure 5. Analyses of testicular tissue in mice using hematoxylin-eosin (H&E) staining. The testicular tissue of mice was stained with H&E to observe pathologic changes in the testes under electron microscopy (n=6 animals per group). In group A: (i) The convoluted tubules are closely arranged, and the boundary between the basal membrane and the stroma is clear and even (arrow ?); (ii) all levels of spermatogenic cells and spermatogenic cells are arranged in order (arrows ?, ?). In group B: (i) Spermatogenic tubules are injured to different degrees, and the seminiferous tubules were sparsely arranged, showing interstitial edema, and their epithelium was separated from the matrix (arrow ?); (ii) the number of Sertoli cells in testes was significantly reduced and arranged loosely. The number of spermatozoa was significantly reduced (arrows ?, ?). 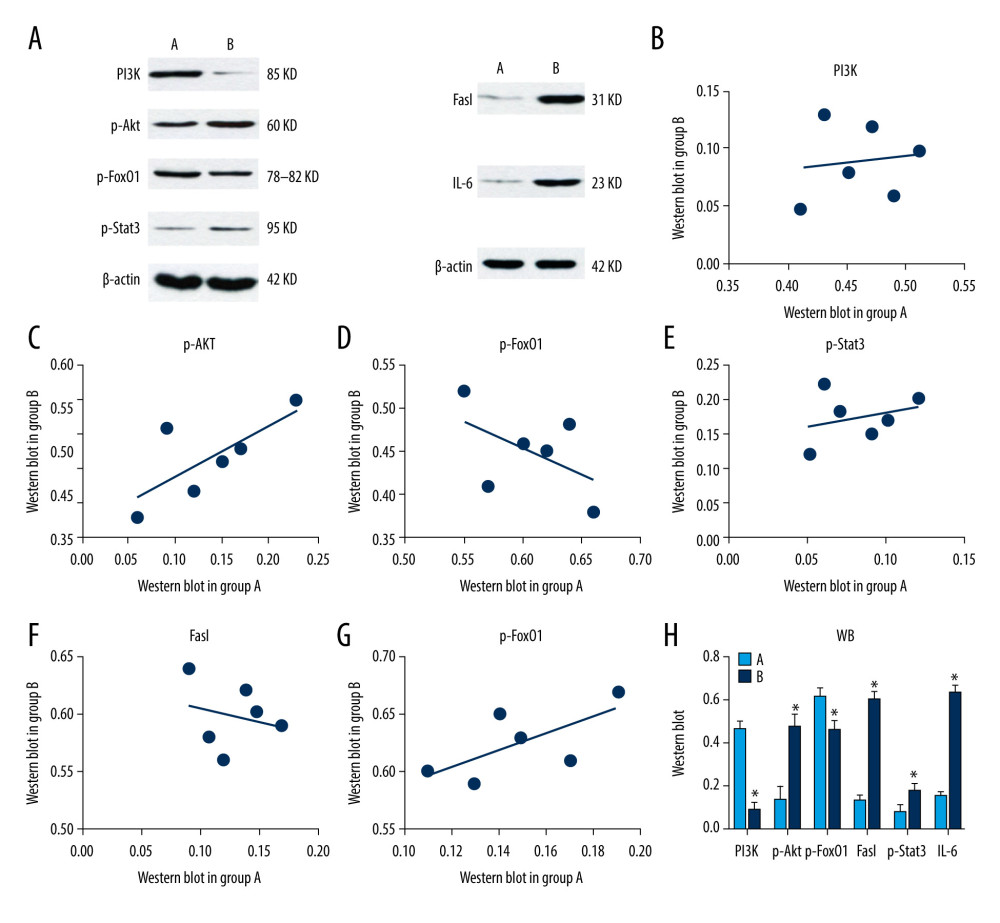 Figure 6. (A) Western blotting showing expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 proteins. β-actin is a loading control. Scatter plot of western blot expression of (B) PI3K, (C) p-Akt, (D) p-FoxO1, (E) FasL, (F) p-Stat3, and (G) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (H) The light blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01.
Figure 6. (A) Western blotting showing expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 proteins. β-actin is a loading control. Scatter plot of western blot expression of (B) PI3K, (C) p-Akt, (D) p-FoxO1, (E) FasL, (F) p-Stat3, and (G) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (H) The light blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01. 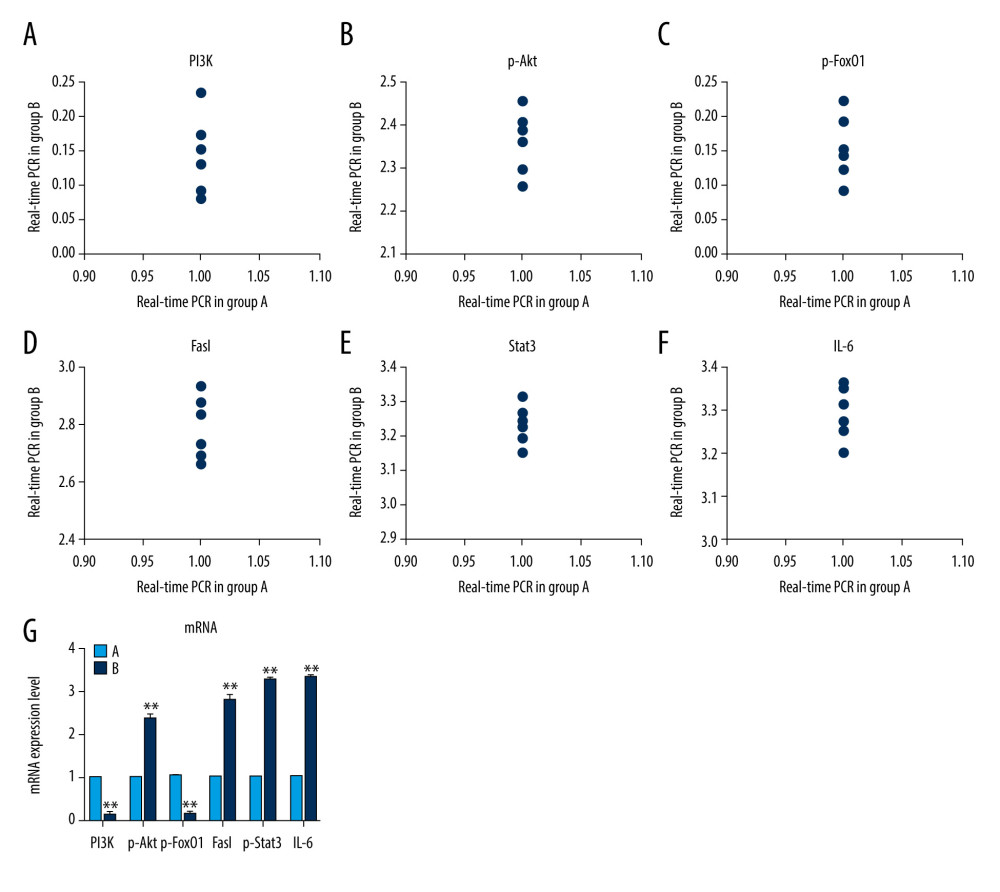 Figure 7. Scatter plot of mRNA expression of (A) PI3K, (B) p-Akt, (C) p-FoxO1, (D) FasL, (E) p-Stat3, and (F) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (G) Expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in mice testes. The light blue bar graph represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01.
Figure 7. Scatter plot of mRNA expression of (A) PI3K, (B) p-Akt, (C) p-FoxO1, (D) FasL, (E) p-Stat3, and (F) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (G) Expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in mice testes. The light blue bar graph represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01. 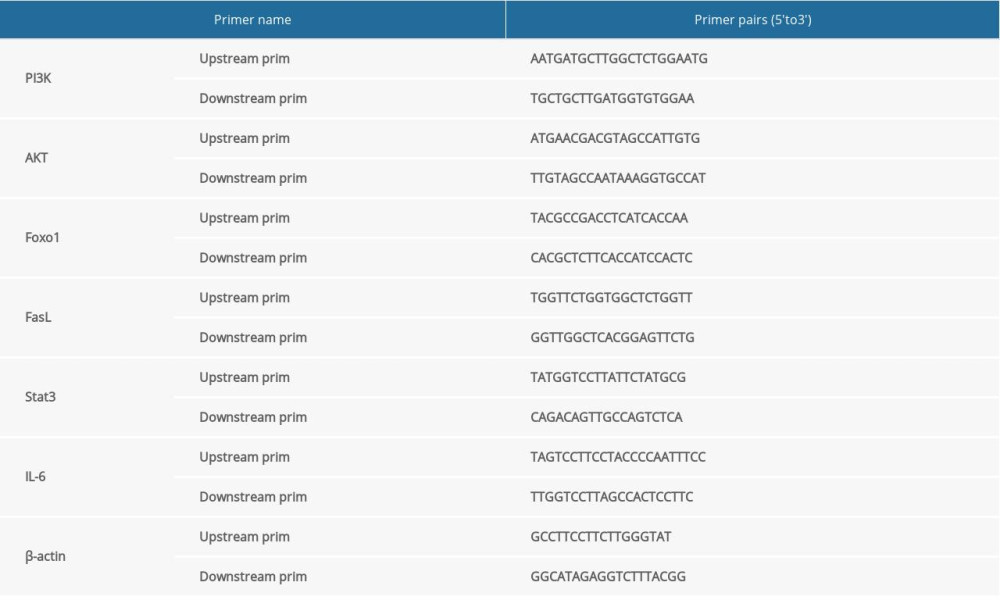
References
1. Zegers-Hochschild F, Adamson GD, de Mouzon J, International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology: Fertil Steril, 2009; 92; 1520-24
2. Agarwal A, Parekh N, Selvam MKP, Male oxidative stress infertility (MOSI): Proposed terminology and clinical practice guidelines for management of idiopathic male infertility: World J Mens Health, 2019; 37(3); 296-312
3. Kolesnikova LI, Kolesnikov SI, Kurashova NACauses and factors of male infertility: Vestn Ross Akad Med Nauk, 2015(5); 579-84 [in Russian]
4. Ge S, Zhao P, Liu X, Necessity to evaluate epigenetic quality of the sperm for assisted reproductive technology: Reprod Sci, 2019; 26(3); 315-22
5. Zhang S, Wu J, Liu XTraditional Chinese Medicine in seminal abnormality: A medication analysis based on data mining: World Chinese Med, 2017; 12(7); 1688-92 [in Chinese]
6. Alves MG, Martins AD, Rato L, Molecular mechanisms beyond glucose transport in diabetes-related male infertility: Biochim Biophys Acta, 2013; 1832(5); 626-35
7. Dias TR, Alves MG, Silva BM, Oliveira PF, Sperm glucose transport and metabolism in diabetic individuals: Mol Cell Endocrinol, 2014; 396(1–2); 37-45
8. Maresch CC, Stute DC, Alves MG, Diabetes-induced hyperglycemia impairs male reproductive function: A systematic review: Hum Reprod Update, 2018; 24(1); 86-105
9. Paramio JM, Jorcano JL, Beyond structure: Do intermediate filaments modulate cell signalling?: Bioessays, 2002; 24(9); 836-44
10. Yan M, Shi Y, Wang Y, Effects of p, p’-DDE on the mRNA and protein expressions of vimentin, N-cadherin and FSHR in rats testes: An in vivo and in vitro study: Environ Toxicol Pharmacol, 2013; 35(3); 486-94
11. Huang YT, van der Hoorn D, Ledahawsky LM, Robust comparison of protein levels across tissues and throughout development using standardized quantitative western blotting: J Vis Exp, 2019(146); 59438
12. Yang P, Gong YJ, Wang YX, Effect modification by apoptosis-related gene polymorphisms on the associations of phthalate exposure with spermatozoa apoptosis and semen quality: Environ Pollut, 2017; 231(Pt 1); 694-702
13. Gao S, Zhang Y, Yang C, NCOA5 haplo-insufficiency results in male mouse infertility through increased IL-6 expression in the epididymis: Sci Rep, 2019; 9(1); 15525
14. Gao Q, Wolfgang MJ, Neschen S, Disruption of neural signal transducer and activator of transcription 3 causes obesity, diabetes, infertility, and thermal dysregulation: Proc Natl Acad Sci USA, 2004; 10(13); 4661-66
15. Piccoli GB, Grassi G, Cabiddu G, Diabetic kidney disease: A syndrome rather than a single disease. The review of diabetic studies: Rev Diabet Stud, 2015; 12(1–2); 87-109
16. Askari H, Rajani SF, Poorebrahim M, A glance at the therapeutic potential of irisin against diseases involving inflammation, oxidative stress, and apoptosis: An introductory review: Pharmacol Res, 2018; 129; 44-55
17. Zhang H, Ge S, He K, FoxO1 inhibits autophagosome-lysosome fusion leading to endothelial autophagic-apoptosis in diabetes: Cardiovasc Res, 2019; 115(14); 2008-20
18. Fu WJ, Wang CM, Quan CY, Effects of bisphenol A on expression of P38 MAPK and caspase 3 in cultured primary Sertoli cells of rats: J Environ Health, 2013; 6; 481-85
19. Zhang PH, Chen DA, Dong L, Inhibitory effect of Qiangjing tablets on the Fas/FasL pathway of cell apoptosis in male SD rats with infertility: J Androl, 2016; 22(03); 246-51
20. Liu Q, Wang JJ, Hu YQBerberine regulates PI3K/AKT/FOXO1/Bim signaling pathway to improve high glucose-induced podocyte injury: Chinese Pharmacol Bull, 2020; 36(3); 329-34 [in Chinese]
21. Zhang L, Liu X, Nie XEffects of salvianolate on TLR4/IL-6/STAT3 pathway in rats with atherosclerosis: Academic Journal of Shanghai University of Traditional Chinese Medicine, 2020; 34(4); 61-65 [in Chinese]
22. Kim DH, Park MH, Lee EK, The roles of FoxOs in modulation of aging by calorie restriction: Biogerontology, 2015; 16; 1-14
23. Cheng , White MF, Targeting Forkhead box O1 from the concept to metabolic diseases: lessons from mouse models: Antioxid Redox Sign, 2011; 14(4); 649-61
24. Wang Q, Wang N, Dong M, GdCl3 reduces hyperglycaemia through Akt/FoxO1-induced suppression of hepatic gluconeogenesis in Type 2 diabetic mice: Clin Sci, 2014; 127(2); 91-100
25. Manning BD, Cantley LC, AKT/PKB signaling: Navigating downstream: Cell, 2007; 129(7); 1261-74
26. Del Giudice PT, Lima SB, Cenedeze MA, Expression of the Fas-ligand gene in ejaculated sperm from adolescents with and without varicocele: J Assist Reprod Genet, 2010; 27(2–3); 103-9
27. Xu YN, Yu CM, Hu LLThe FAS/FASL single nucleotide polymorphism in Southern China population: Guangdong Medical Journal, 2017; 38(9); 1333-37 [in Chinese]
28. Cao Q, Ge X, Wang MChanges of serum 25-hydroxyvitamin D3 in patients with type 2 diabetic nephropathy and its correlation with renal function indexes and inflammatory factors: Chinese Journal of Difficult and Complicated Cases, 2015; 14(12); 1245-48 [in Chinese]
29. Aghazadeh S, Yazdanparast R, Mycophenolic acid potentiates HER2-overexpressing SKBR3 breast cancer cell line to induce apoptosis: Involvement of AKT/FOXO1 and JAK2/STAT3 pathways: Apoptosis, 2016; 21(11); 1302-14
Figures
Figure 1. Overall process based on biological network research and animal experiments.Figure 2. (A) Intersection of targets of diabetes mellitus and male infertility; (B) Diabetes mellitus-male infertility targets network built by Cytoscape (3.7.1); (C) protein–protein interaction (PPI) network built by Cytoscape (3.7.1); (D) PPI network processed by Cytoscape (3.7.1) plug-in (cytohubba).Figure 3. (A–D) Analyses of pathway enrichment using Gene Ontology and Kyoto Encyclopedia of Genes and Genomes databases. The Y axis represents the gene name and the X axis represents the richness factor. The size of the node is proportional to the number of genes. The node color is proportional to the P value.Figure 4. (A) Blood glucose of mice from groups A (control) and B (diabetic). (C, E) Sperm counts and total sperm viability in the 2 groups. (B, D, F) Scatter plot of blood glucose, sperm count and total sperm viability in the 2 groups of mice. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A, * P<0.05.Figure 5. Analyses of testicular tissue in mice using hematoxylin-eosin (H&E) staining. The testicular tissue of mice was stained with H&E to observe pathologic changes in the testes under electron microscopy (n=6 animals per group). In group A: (i) The convoluted tubules are closely arranged, and the boundary between the basal membrane and the stroma is clear and even (arrow ?); (ii) all levels of spermatogenic cells and spermatogenic cells are arranged in order (arrows ?, ?). In group B: (i) Spermatogenic tubules are injured to different degrees, and the seminiferous tubules were sparsely arranged, showing interstitial edema, and their epithelium was separated from the matrix (arrow ?); (ii) the number of Sertoli cells in testes was significantly reduced and arranged loosely. The number of spermatozoa was significantly reduced (arrows ?, ?).Figure 6. (A) Western blotting showing expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 proteins. β-actin is a loading control. Scatter plot of western blot expression of (B) PI3K, (C) p-Akt, (D) p-FoxO1, (E) FasL, (F) p-Stat3, and (G) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (H) The light blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01.Figure 7. Scatter plot of mRNA expression of (A) PI3K, (B) p-Akt, (C) p-FoxO1, (D) FasL, (E) p-Stat3, and (F) IL-6 protein in the 2 groups of mice: group A (control) and group B (diabetic). (G) Expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in mice testes. The light blue bar graph represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group A. The deep blue bar chart represents expression of the mRNA of PI3K, p-Akt, p-FoxO1, FasL, p-Stat3, and IL-6 in group B. Values are the mean±SEM (n=6 animals per group). The t test was used to compare group B with group A. * P<0.05, ** P<0.01. In Press
Clinical Research
Effects of Single-Bout Endurance Exercise Intensity on Peripheral Neurotrophic Factors in Patients With Isc...Med Sci Monit In Press; DOI: 10.12659/MSM.952089
Review article
Anisodus tanguticus in Cancer Research: A Review of Traditional Use, Phytochemistry, Extraction Methods, an...Med Sci Monit In Press; DOI: 10.12659/MSM.952999
Clinical Research
Nasal Mucociliary Clearance and Its Relationship With Disease Severity in Patients With Multiple SclerosisMed Sci Monit In Press; DOI: 10.12659/MSM.952850
Clinical Research
Modified Thoracoabdominal Nerves Block Through the Perichondrial Approach vs Subcostal Transversus Abdomini...Med Sci Monit In Press; DOI: 10.12659/MSM.953976
Most Viewed Current Articles
17 Jan 2024 : Review article 14,176,570
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
13 Nov 2021 : Clinical Research 3,762,188
Acceptance of COVID-19 Vaccination and Its Associated Factors Among Cancer Patients Attending the Oncology ...DOI :10.12659/MSM.932788
Med Sci Monit 2021; 27:e932788
14 Dec 2022 : Clinical Research 2,466,310
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research 708,927
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387